Экспериментальная психология
2025. Том 18. № 1. С. 138–154
doi:10.17759/exppsy.2025180109
ISSN: 2072-7593 / 2311-7036 (online)
Анализ мозговой активности при конфигурационном научении с помощью магнитоэнцефалографии
Аннотация
Контекст и актуальность. Работа посвящена исследованию кодирования мозгом комплексных стимулов при конфигурационном ассоциативном научении у человека. Поведение, которое основано на восприятии комплексных сигналов, обеспечивает высокую адаптивность человеческой деятельности. При этом на настоящий момент знания об участии коры больших полушарий в связывании стимульных элементов в воспринимаемую целостную конфигурацию остаются неполными и противоречивыми. Методы и материалы. Мы использовали четыре элементных разной модальности (два зрительных и два слуховых) и два комплексных мультимодальных стимула, составленные из тех же элементных стимулов. Два стимула (один комплексный и один элементный) сочетали с отрицательным подкреплением (электрокожным раздражением). Задача испытуемого состояла в нажатии кнопки в случае возникновения у него ожидания электрокожного раздражения после предъявления каждого стимула: стимулы предъявлялись в псевдослучайном порядке. В исследовании приняли участие 29 добровольных участников. Результаты исследования показали, что подкрепление комплексного стимула сопровождается значимым увеличением мощности тета-осцилляций в ответ на этот стимул. Кроме того, выявлено, что кодирование конфигурационной ассоциации вовлекает тета-осцилляции в большей степени в сравнении с элементной ассоциацией. Эти эффекты выявлены в локализациях, указывающих на префронтальную кору, левые дорсолатеральные фронтальные области, правые височные области и теменно-затылочные области. Выводы. Мы предполагаем, что этот феномен не только является следствием вовлечения гиппокампа в кодирование комплексного стимула, но и свидетельствует об активном взаимодействии между гиппокампом и ассоциативными областями новой коры в процессе научения.
Общая информация
Ключевые слова: научение, ключевые стимулы, осцилляции, магнитоэнцефалография (МЭГ)
Рубрика издания: Психофизиология
Тип материала: научная статья
DOI: https://doi.org/10.17759/exppsy.2025180109
Финансирование. Исследование выполнено за счет гранта Российского научного фонда № 23-78-00010.
Благодарности. Исследование выполнено на Уникальной научной установке «Центр нейрокогнитивных исследований (МЭГ-центр)» МГППУ. Исследование выполнено при поддержке Междисциплинарной научно-образовательной школы Московского государственного университета «Мозг, когнитивные системы, искусственный интеллект».
Получена: 06.08.2024
Принята в печать:
Для цитаты: Денисова Е.В., Позняк Л.А., Пульцина К.И., Третьякова В.Д., Чернышев Б.В. Анализ мозговой активности при конфигурационном научении с помощью магнитоэнцефалографии // Экспериментальная психология. 2025. Том 18. № 1. С. 138–154. DOI: 10.17759/exppsy.2025180109
Полный текст
Введение
Ассоциативное научение может принимать две качественно различные формы в зависимости от уровня репрезентации условного стимула в восприятии субъекта. Элементный механизм формирования ассоциаций подразумевает, что научение происходит путем ассоциации подкрепления с отдельными элементами или признаками условных стимулов (например, высота звукового тона или цвет зрительного сигнала). В отличие от элементного, конфигурационный механизм предполагает, что научение вырабатывается в ответ на общую структуру или конфигурацию стимульного массива в целом (Rudy, Sutherland, 1995; Чернышев, Ушаков, Позняк, 2024). У человека и животных с высокоразвитым мозгом имеется тенденция к формированию ассоциаций именно на целостные стимульные конфигурации (Kimchi, 1994; Navon, 1977), что обеспечивает более высокую адаптивность поведения и даже считается наиболее ранним предвестником возникновения сознания (Bronfman, Ginsburg, Jablonka, 1954; Feinberg, Mallatt, 2016; Ginsburg, Jablonka, 2019; Razran, 1971). Однако нейрокогнитивные механизмы конфигурационного научения до сих пор остаются малоизученными.
Конфигурационное научение, в отличие от элементного, критически зависит от гиппокампа (Rudy, Sutherland, 1995; Чернышев, Ушаков, Позняк, 2024): считается, что именно гиппокамп обеспечивает возможность создания целостных конфигурационных представлений комплексных стимулов на основе временных и пространственных взаимосвязей между стимульными элементами (Rudy, Huff, Matus-Amat, 2004; Rudy, Sutherland, 1995; Sutherland, Rudy, 1989). Вероятно, конфигурационные репрезентации после кодирования в гиппокампе транслируются в другие кортикальные и подкорковые структуры (например, медиальную префронтальную кору и миндалину), где, собственно, происходит само научение — ассоциация поступившей стимульной конфигурации со значимым подкреплением. Вовлечение гиппокампа в конфигурационное научение было выявлено в ряде современных работ на человеке с применением функциональной магниторезонансной томографии (Baeuchl et al., 2015; Duncan et al., 2018; Stout et al., 2019). Если же у человека функция гиппокампа нарушена, то тогда сложная информация может быть представлена в его мозге лишь на уровне элементных ассоциаций (Maren, Aharonov, Fanselow, 1997; Maren, Phan, Liberzon, 2013).
Известной характерной особенностью электрофизиологии гиппокампа являются осцилляции в тета-диапазоне, которые являются важным организующим механизмом, обеспечивающим функции гиппокампа при многих видах деятельности животного и человека, требующих активного вовлечения в выполнение задачи (Buzsáki, McKenzie, Davachi, 2022; Lisman, Buzsaki, 2008). При этом кодирование входящей информации и формирование памяти осуществляется благодаря интенсивному взаимодействию в тета-диапазоне между гиппокампом и неокортексом (Karakaş, 2020; Miller, 2013). В последнее время в литературе уделяется значительное внимание взаимодействию между гиппокампом и новой корой в отношении кодирования и удержания памяти (Jensen, Hennequin, Mattar, 2024; Joensen et al., 2023; Nardin et al., 2023).
Однако результаты целого ряда исследований на животных с проведением регистрации мощности гиппокампальных тета-осцилляций при конфигурационном научении (Sakimoto et al., 2013; Sakimoto et al., 2013; Sakimoto, Sakata, 2015) приводят к неоднозначным выводам — обнаруживается как повышение, так и понижение мощности тета-осцилляций.
В исследованиях на людях было показано, что конфигурационное научение специфически задействует осцилляции в тета-диапазоне (Cashdollar et al., 2009; Fuentemilla et al., 2010; Poch et al., 2011). В частности, выявлена сеть тета-осцилляций, вовлеченная в удержание конфигурационных репрезентаций и объединяющая затылочные и височные области; важно, что у пациентов с пораженным гиппокампом данная сеть не выявлялась (Cashdollar et al., 2009).
В работе Олсена (Olsen) и др. (Olsen et al., 2013) было показано, что мощность тета-осцилляций в гиппокампе и медиальной префронтальной коре увеличивалась в периоды кодирования и удержания в памяти целостной конфигурации зрительных стимулов. Также были выявлены эффекты в правой латеральной префронтальной коре и внутритеменной борозде в случае изменения конфигурации — по сравнению с сохранением расположения элементов зрительного стимула. Память на конфигурационные отношения была положительно связана с повышением мощности тета-осцилляций в гиппокампе.
Наиболее распространенным подходом к изучению конфигурационного научения у человека является обстановочное (контекстное) научение, при котором отдельные элементы сочетаются в целостное представление о конкретном месте в окружающей среде, где происходит какое-либо значимое событие (Baeuchl et al., 2015; Stout et al., 2019; Stout et al., 2018). В этих работах использовались зрительные предъявления изображений комнат, в которых размещены разнообразные элементы обстановки (столы, стулья, диваны, торшеры, и т. п.), либо использовались пространственные комбинации более сложных зрительных элементов как условный стимул. Методологическим недостатком выполненных ранее работ, посвященных исследованию конфигурационного научения на человеке, является применение стимулов лишь в пределах одной зрительной модальности, причем в весьма специфическом контексте обстановочного научения. Однако именно межмодальные конфигурационные отношения между стимулами позволили выявить специфическую активацию гиппокампа при конфигурационном научении, в отличие от выработки реакции на его элементные компоненты комплексного стимула, используемые по отдельности (Ивашкина и др., 2020).
Цель настоящего магнитоэнцефалографического исследования состояла в том, чтобы выявить специфические отличия в мощности тета-осцилляций, характеризующие формирование ассоциации именно при подкреплении комплексных стимулов — в сравнении с подкреплением элементных, а также в сравнении с комплексными стимулами в отсутствие подкрепления. Мы исходили из предположения о том, что если комплексные стимулы вовлекаются в ассоциацию с подкреплением, то мозг конструирует из репрезентаций сочетающихся стимульных элементов целостную конфигурацию, которая и вовлекается в ассоциацию. Этот процесс требует участия гиппокампа и большего взаимодействия гиппокампа с корой больших полушарий, проявляющегося в усилении мощности тета-осцилляций в коре больших полушарий. В случае, если комплексный стимул не вовлекается в ассоциацию, а также в случае, если стимул является элементным, указанные процессы должны быть менее выражены.
В нашем исследовании конфигурационного научения мы впервые использовали дизайн с мультимодальными конфигурациями, что позволило отойти от более узкой проблемы контекстного научения и исследовать формирование конфигураций как таковых. Сбалансированный дизайн позволил сопоставить активность мозга при формировании ассоциаций на комплексные стимулы и на элементные стимулы, а также сравнить формирование ассоциаций на комплексный стимул с ситуацией, когда комплексный стимул является дифференцировочным и не ассоциируется с подкреплением. Результаты анализа указывают на статистически значимое увеличение мощности тета-осцилляций при научении на подкрепляемый комплексный стимул в сравнении с остальными типами стимулов.
Материалы и методы
В эксперименте приняли участие 29 добровольных участников в возрасте от 18 до 42 лет (средний возраст — 28 лет, 11 мужчин), обязательными требованиями выступала праворукость и владение русским языком как родным. Все участники имели нормальный слух и не страдали от неврологических или психиатрических расстройств.
Для изучения элементного и конфигурационного научения участникам предъявляли четыре элементных стимула (два зрительных и два слуховых) и два комплексных стимула, представляющих собой межмодальные сочетания тех же зрительных и слуховых стимулов. Таким образом, совокупно процедура включала в себя предъявление 6 разных стимулов (рис. 1, таблица).
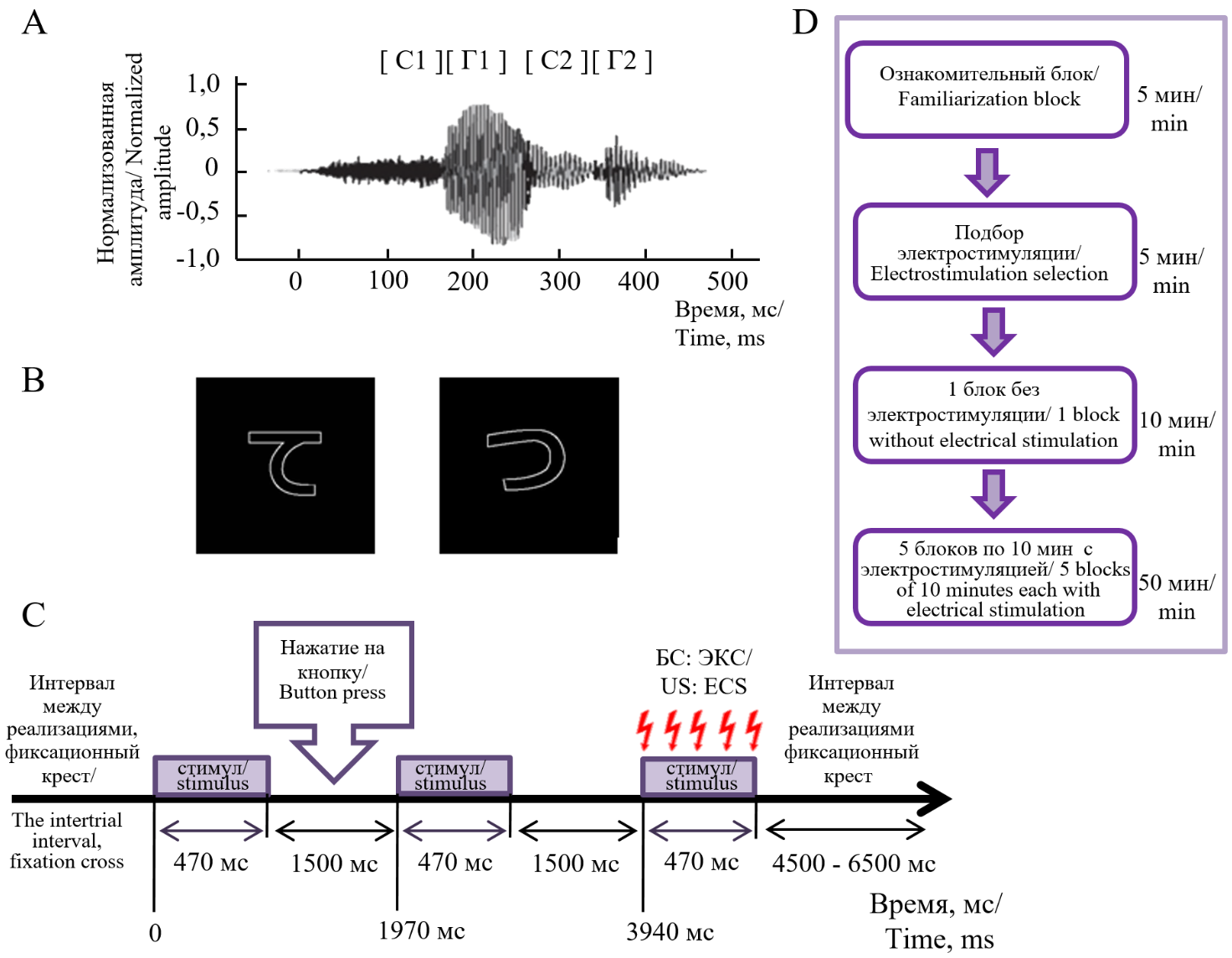
Рис. 1. Стимулы и экспериментальный дизайн: А — Пример построение псевдослова;
B — Изображение зрительных стимулов; C — Временная организация реализации эксперимента (БС — безусловный стимул, ЭКС — электрокожная стимуляция); Ноль на графиках соответствует моменту начала предъявления стимула; D — Схема последовательности блоков
Fig. 1. Stimuli and experimental design: A — Example of pseudoword construction; B — Visual stimuli images; C — Temporal organization of the experimental procedure (US — unconditioned stimulus, ECS — electrocutaneous stimulation); Zero on the graphs corresponds to the moment of stimulus onset; D — Diagram of the block sequence
Таблица / Table
Стимулы и контрбалансировка по группам
Stimuli and counterbalancing across groups
|
Набор стимулов / Set of stimuli |
Тип стимула / Stimulus type |
Стимулы/Stimuli |
Контрбалансировочные группы / Counterbalancing groups |
|||
|
1 |
2 |
3 |
4 |
|||
|
1 |
Комплексный/Complex |
Слуховой стимул С1 / и Зрительный стимул Зр1 / |
̶ |
̶ |
+ |
+ |
|
Элементный/Elemental |
Слуховой стимул С1 / |
+ |
̶ |
̶ |
̶ |
|
|
Элементный/Elemental |
Зрительный стимул Зр1 / |
̶ |
+ |
̶ |
̶ |
|
|
2 |
Комплексный/ Complex |
Слуховой стимул С2 / и Зрительный стимул Зр2 / |
+ |
+ |
̶ |
̶ |
|
Элементный/ Elemental |
Слуховой стимул С2 / |
̶ |
̶ |
+ |
̶ |
|
|
Элементный/ Elemental |
Зрительный стимул Зр2 / |
̶ |
̶ |
̶ |
+ |
|
Примечание: стимулы, сочетавшиеся с электрокожной стимуляцией, обозначены символом «+», без стимуляции — символом «–».
Note: Stimuli paired with electrocutaneous stimulation are marked with symbol «+», while those without stimulation are marked with symbol «–».
Слуховые стимулы являлись псевдословами, акустические и фонетические свойства которых были уравновешены (Razorenova et al., 2020) (рис. 1А). Использовали слоги, которые состояли из одной согласной и одной гласной (CГ), и составили из них два двуслоговых (C1Г1C2Г2) бессмысленных псевдослова: «хи́ча» и «хи́шу». Зрительные стимулы представляли собой изображения двух иероглифов из японской слоговой азбуки Хирагана, выполненные белым контуром на черном фоне (рис. 1B). Зрительные стимулы также были выровнены по общей яркости и длине линий. Длительность предъявления каждого из зрительных, слуховых и комплексных стимулов составляла 470 мс. В каждой реализации стимулы предъявляли по три раза, с интервалом между предъявлениями 1500 мс. Суммарная длительность реализации составляла 4410 мс. Интервал между реализациями составлял 4500—6500 мс (рис. 1C).
В начале эксперимента в ознакомительном блоке участникам зачитывали инструкцию, знакомили их со стимулами и обучали правильно и своевременно нажимать на кнопки (см. ниже), при этом применяли дополнительные наборы стимулов, которые не использовались в основном эксперименте.
Силу тока для электрокожной стимуляции (ЭКС) подбирали исходя из индивидуальной болевой чувствительности каждого участника с целью минимизации дискомфорта. ЭКС осуществляли с помощью изолирующего электростимулятора Digitimer DS7A (Digitimer, Welwyn Garden City, UK) через одноразовые гелевые Ag/Cl-электроды, которые прикрепляли ко вторым фалангам мизинца и безымянного пальца правой руки.
Основная часть эксперимента состояла из шести блоков (рис. 1D). В начальном экспериментальном блоке происходило «пассивное» предъявление элементных и комплексных стимулов, без сочетания с электрокожной стимуляцией. В последующих пяти блоках производили сочетание двух из шести стимулов (одного элементного и одного комплексного, см. ниже) с ЭКС (рис. 1C), а остальные четыре не сопровождались ЭКС. Какие именно два из шести стимулов сочетались с ЭКС, определялось распределением участников по контрбалансировочным группам (таблица), которые мы использовали для минимизации возможных эффектов, связанных с физическими различиями между стимулами. Каждого участника до эксперимента назначали в одну из контрабалансировочных групп квазислучайным образом, при этом обеспечивая однородность состава групп участников по полу и возрасту.
В каждом блоке стимулы предъявляли участникам вперемешку в квазислучайном порядке. Каждый блок суммарно содержал последовательность из 60 реализаций (по 10 раз для каждого стимула).
Согласно процедуре эксперимента, во всех реализациях участников просили предсказывать ожидание ЭКС, нажимая на одну из трех кнопок пальцами правой руки, сразу после первого из трех предъявлений стимула (рис. 1C). Каждая кнопка соответствовала определенному ответу: левая кнопка (указательный палец) — стимул не сопровождается электростимуляцией; средняя кнопка (средний палец) — участник сомневается и не знает, сопровождается ли стимул электростимуляцией или не сопровождается; правая кнопка (безымянный палец) — стимул сопровождается электростимуляцией. В начальном «пассивном» блоке участников просили всегда нажимать только левую кнопку «не будет».
Поскольку нас интересовали устойчивые эффекты при реализации только что сформированной ассоциации, то в анализ данных МЭГ были включены только последние четыре активных блока. «Пассивный» блок и первый активный блок исключили из анализа, так как на этом этапе ассоциативная память еще не сформировалась.
Регистрацию активности мозга проводили с помощью магнитоэнцефалографической установки «Elekta Neuromag Vector View» (Хельсинки, Финляндия). Сигнал МЭГ (306 каналов, включая 204 планарных градиентометров) записывали с использованием встроенных фильтров с полосой пропускания 0,03—330 Гц, частота дискретизации составляла 1000 Гц. Сигнал МЭГ далее обрабатывали с применением программного обеспечения MNE-Python toolbox v.1.7.0 (Gramfort et al., 2014). Для дальнейшего анализа использовали сигнал, полученный от 204 планарных градиентометров, причем ортогональные градиентометры объединяли, вычисляя среднеквадратичное среднее значение в каждой паре. В итоге анализ проводили на 102 объединенных сенсорах.
Магнитоэнцефалографические данные очищали от артефактов, связанных с сердцебиением и движениями глаз, используя анализ независимых компонент (Independent Component Analysis, ICA). Для последующего анализа данные были разделены на эпохи — от -2 до 4 с относительно момента начала первого предъявления стимула. Далее извлекали из исходного сигнала мощность осцилляций в пределах интересующего нас тета-диапазона, который составлял от 4 до 8 Гц, используя функцию «mne.time_frequency.tfr_multitaper» (из библиотеки MNE-Python), которая основана на методе частотно-временных представлений сигналов с использованием функции Слепиана («multitapers», discrete prolate spheroidal sequences). Использовали скользящее окно длительностью 0,5 с, (т.е. –2—4 цикла осцилляций), с частотами от 4 от 8 Гц, с шагом 1 Гц. Полученные данные по узким частотным полосам суммировали, чтобы получить суммарную мощность в тета-диапазоне. Анализ проводили в интервале от 0,1 до 0,8 с после предъявления первого стимула; именно в этом интервале происходят основные события в активности мозга, вызванные стимулом, включая его анализ и кодирование (Luck, 2014; Polich, 2007). Коррекции базовой линии осуществлялась на временном интервале перед первым предъявлением стимула, начиная с –1 с и заканчивая –0,4 с относительно начала предъявления.
Для статистической обработки использовали среду программирования R версия 4.3.3 (R Core Team, 2021). Для нахождения значимых сенсоров, отражающих влияние типа стимула и условия подкрепления на мощность тета-осцилляций, мы применили линейные смешанные модели (Linear Mixed-Effects Model, LMM) с поправкой на множественные сравнения с помощью метода ожидаемой доли ложных отклонений (false discovery rate, FDR) на число сенсоров анализа. Для анализа использовали функции «lmer» и «mul.fdrcorrection». Для оценки зависимости тета-мощности от типа стимула в модель LMM в качестве фиксированных эффектов включили тип стимула (Тип_стимула: элементный и комплексный) и условие подкрепления (Условие_подкрепляемости: подкрепляемые и неподкрепляемые стимулы) и их взаимодействие, различие между участниками (участник) было включено в качестве случайного эффекта:
Тета-мощность ~ Тип_стимула * Условие_подкрепляемости + (1| участник).
Далее мы усреднили значения мощности тета-осцилляций по всем сенсорам, в которых на предыдущем этапе анализа было обнаружено достоверное взаимодействие факторов: Тип_стимула * Условие_подкрепляемости (по порогу p < 0,05, с FDR-коррекцией), а также дополнительно по топографическим кластерам в пределах множества значимых сенсоров. На данном этапе анализа мы применили аналогичную LMM-модель, но для фактора «Тип_стимула» использовали три уровня: элементный зрительный, элементный слуховой и комплексный. Для апостериорного попарного сравнения групп после LMM применяли тест Тьюки (Tukey's HSD test). Для построения временных кривых данные усредняли в последовательных временных точках с шагом 0,05 с.
Результаты
В процессе ассоциативного научения участники успешно освоили задачу различения стимулов с подкреплением и без — как комплексных, так и элементных. Они в среднем достигли более чем 75% правильных ответов уже в первом активном блоке, а начиная со второго активного блока и далее показатели держались на плато около 90% (рис. 2).
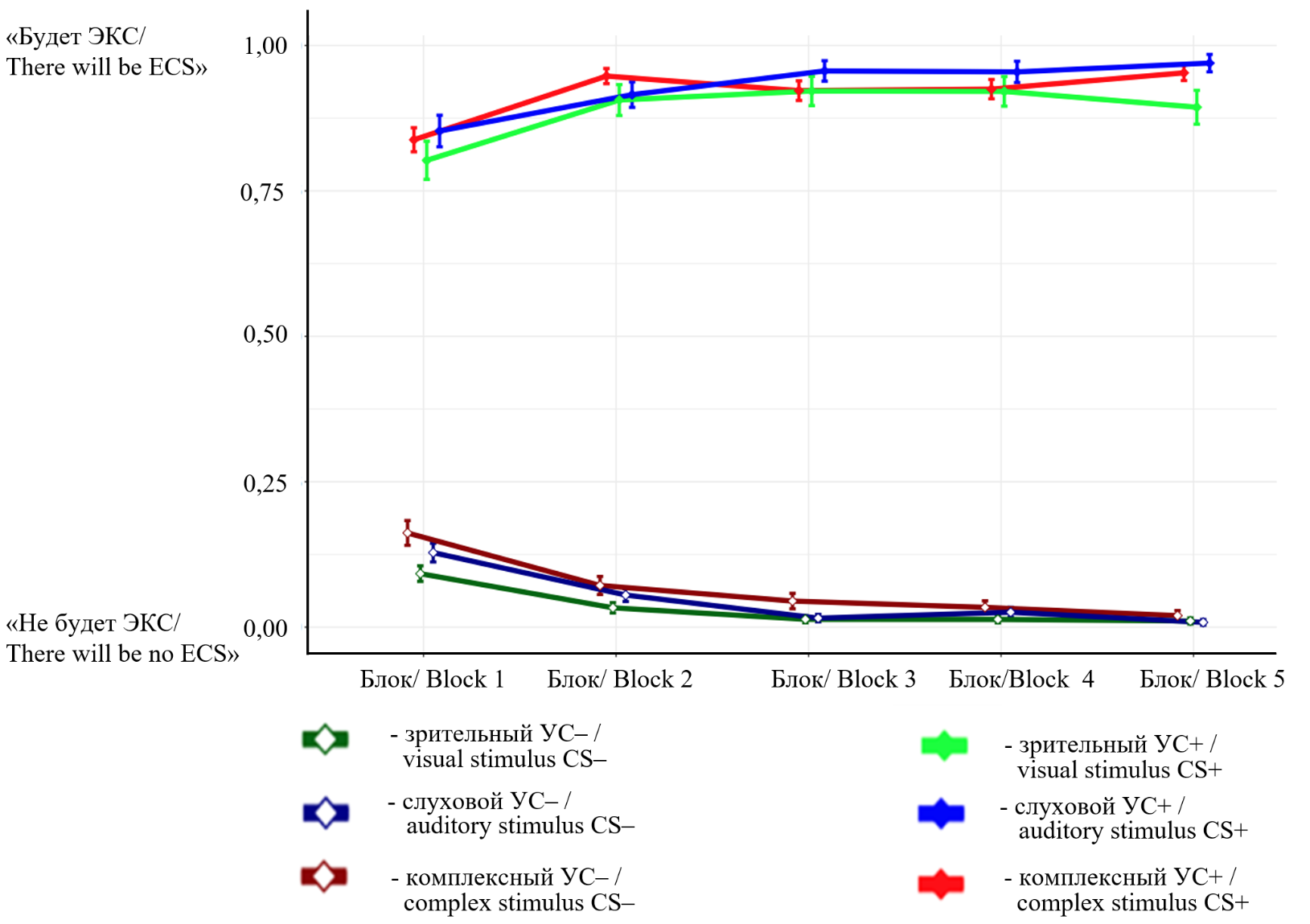
Рис. 2. Усредненные кривые научения по блокам. Доли ответов участников: «0» — стимул является неподкрепляемым («электростимуляцией не сопровождается»); «1» — стимул подкрепляемый («сопровождается электростимуляцией»); «УС» — условный стимул; «УС+» и «УС–» — подкрепляемый и неподкрепляемый условные стимулы соответственно
Fig. 2. Averaged learning curves across blocks. Participants' response ratios: "0" — the stimulus is non-reinforced ("not accompanied by electrostimulation"); "1" — the stimulus is reinforced ("accompanied by electrostimulation"); "CS" — conditioned stimulus; "CS+" and "CS–" — reinforced and non-reinforced conditioned stimuli, respectively
На начальных этапах анализа мы поставили задачу выявить различия между мозговыми ответами на комплексные и элементные стимулы, исключив влияние специфичности мозговых ответов, обусловленных модальностью стимулов. Для этого мы усреднили данные для слуховых и зрительных элементных стимулов, рассматривая их как одно условие с элементными стимулами, противопоставляемое условию с комплексными стимулами (отдельно для подкрепляемых и неподкрепляемых).
Для анализа пространственного распределения мощности тета-активности мы построили топографические карты мощности на временном интервале от 0,1 до 0,8 с относительно начала первого предъявления стимулов (рис. 3). Как видно на рисунке, более значительная синхронизация возникает при кодировании подкрепляемого комплексного стимула по сравнению с неподкрепляемым, а также подкрепляемого комплексного в сравнении с подкрепляемым элементным. Вторая разница (рис. 3C) показала взаимодействие между стимулами, продемонстрировав, что различие в тета-активности более выражено при комплексном подкрепляемом стимуле по сравнению с элементными стимулами в обоих условиях (как подкрепляемом, так и неподкрепляемом), а также с комплексным неподкрепляемым стимулом.
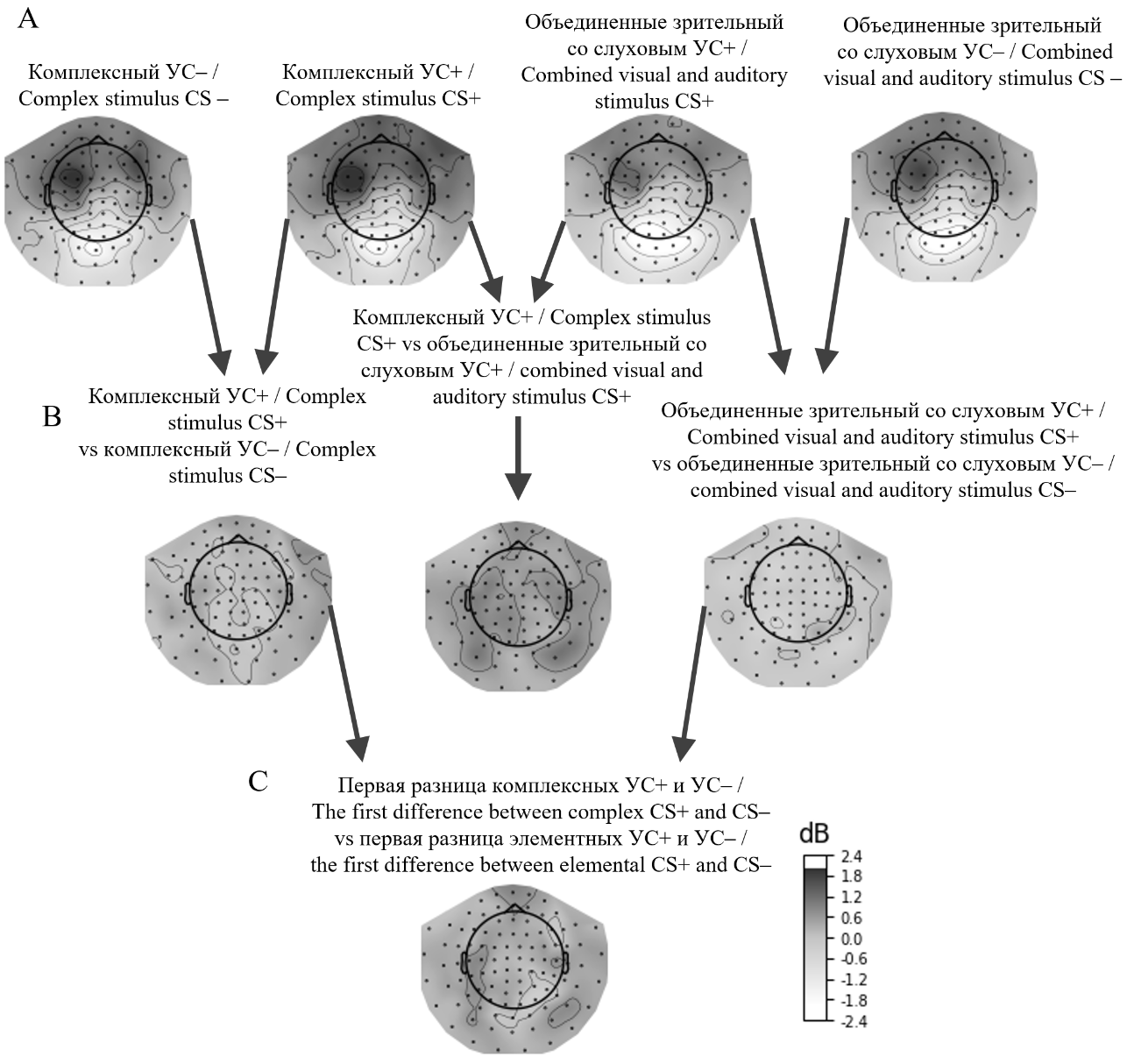
Рис. 3. Карты топографии пространственного распределения разницы мощности тета-активности в ответ на элементные и комплексные стимулы на временном интервале от 0,1 до 0,8 с после начала предъявления стимула: «УС» — условный стимул, «УС+» и «УС–» — подкрепляемый и неподкрепляемый условные стимулы соответственно
Fig. 3. Topographic maps of the spatial distribution of theta activity power difference in response to elemental and complex stimuli within the time interval from 0,1 to 0,8 s after stimulus onset: "CS" — conditioned stimulus, "CS+" and "CS–" — reinforced and non-reinforced conditioned stimuli, respectively
Далее, чтобы доказать существование указанных выше эффектов и изучить их топографию, мы провели статистический анализ с помощью линейных моделей со смешанными эффектами (LMM), с факторами «Тип_стимула» (элементные и комплексные), «Условие_подкрепляемости» (подкрепляемые и неподкрепляемые стимулы) и их взаимодействием. Анализ проводили независимо для каждого из 102 сенсоров, после чего применяли поправку FDR на множественные сравнения на число сенсоров (рис. 4А).
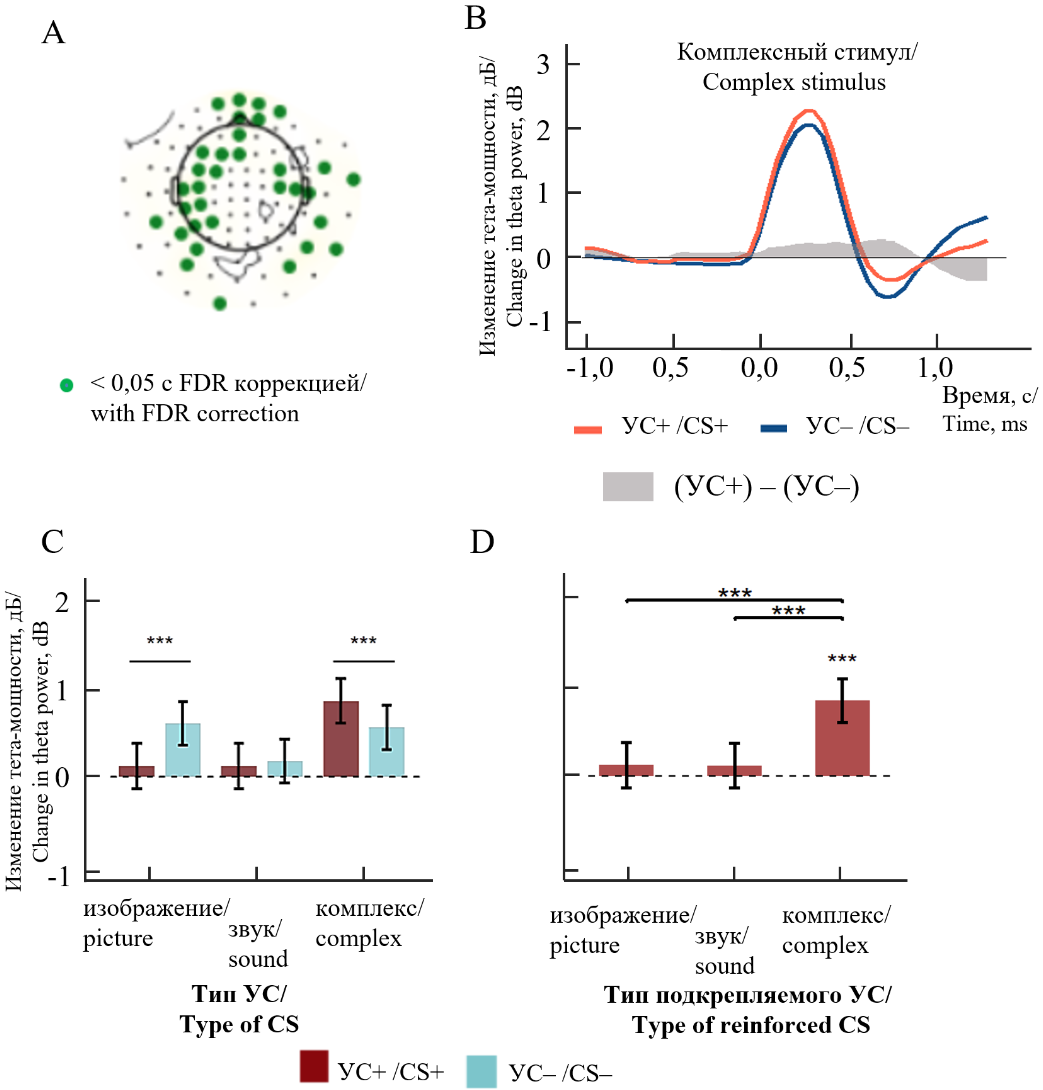
Рис. 4. Анализ мощности тета-осцилляций на усреднении по всем значимым сенсорам: А — достоверные взаимодействия факторов между типами стимулов и подкрепляемостью «Тип стимула*Условие подкрепляемости»; зеленым цветом обозначены статистически значимые сенсоры (p < 0,05); B — временные кривые динамики мощности тета-осцилляций для подкрепляемого и неподкрепляемого комплексного стимула; C — мощность тета-осцилляций в зависимости от типа условного стимула; D — подкрепляемости.
«#» — р < 0,10; «*» — p < 0,05; «**» — p < 0,01; «***» — p < 0,001 в соответствии с тестом Тьюки; «УС» — условный стимул; «УС+» и «УС–» — подкрепляемый и неподкрепляемый условные стимулы соответственно
Fig. 4. Analysis of theta oscillation power averaged across all significant sensors: A — significant interactions between factors of stimulus types and reinforcement conditions "Stimulus Type*Reinforcement Condition"; statistically significant sensors (p < 0,05) are marked in green; B — Time curves of theta oscillation power dynamics for reinforced and non-reinforced complex stimuli; C — Theta oscillation power depending on the type of conditioned stimulus; D — reinforcement condition.
“#” — p < 0,10; “*” — p < 0,05; “**” — p < 0,01; “***” — p < 0,001 according to the Tukey test; “CS” — conditioned stimulus; “CS+” and “CS–” — reinforced and non-reinforced conditioned stimuli, respectively
Нас прежде всего интересовал дифференциальный эффект подкрепления на комплексные стимулы в сравнении с элементными — поскольку согласно нашему предположению, когда комплексные стимулы вовлекаются в ассоциацию с подкреплением, то мозг конструирует из репрезентаций стимульных элементов целостную конфигурацию, которая и вовлекается в ассоциацию. Именно при этом можно ожидать усиленного участия гиппокампа и большего взаимодействия гиппокампа с корой больших полушарий, проявляющегося в усилении мощности тета-осцилляций в коре больших полушарий. Если же комплексный стимул не вовлекается в ассоциацию, а также в случае если стимул является элементным, то можно ожидать значимо более слабой выраженности тета-осцилляций. Соответственно, для дальнейшего анализа мы выбрали все значимые сенсоры по взаимодействию «Тип_стимула * Условие_подкрепляемости» (по порогу p < 0,05, с учетом поправки FDR); на рис. 4A показано их топографическое распределение.
Далее мы перешли к следующему этапу для более детального анализа эффектов, выявленных выше. На этом этапе мы уже не объединяли данные по элементным слуховым и зрительным стимулам, рассматривая теперь их как отдельные уровни фактора «Тип_стимула». Мощность тета-осцилляций усреднили между достоверными сенсорами и снова применили линейную модель со смешанными эффектами, аналогичную описанной выше, с факторами «Тип_стимула» (элементные зрительные, элементные слуховые и комплексные), «Условие_подкрепляемости» (подкрепляемые и неподкрепляемые стимулы) и их взаимодействием. Фактор «Тип_стимула» и взаимодействие факторов «Тип_стимула * Условие_подкрепляемости» находятся на высоком уровне достоверности (F = 48; p < 0,001 и F = 19,9; p < 0,001 соответственно).
Далее, используя построенную LMM-модель, мы получили возможность применить апостериорный критерий Тьюки и сравнить мощности тета-осцилляций между условиями попарно (рис. 4C). Как видно на рисунке, выявлено статистически значимое увеличение тета-мощности на подкрепляемый комплексный стимул по сравнению с неподкрепляемым комплексным (рис. 4C), а для элементных стимулов данного эффекта либо не обнаруживалось (слуховые стимулы), либо он имел обратный знак (зрительные стимулы). Также мы оценили попарные различия между элементными и комплексными стимулами, при условии их подкрепления — т. е. придания им значимости и вовлечения в ассоциацию (рис. 4D). Как видно на рисунке, реакция на комплексный подкрепляемый стимул значимо сильнее, чем для элементных стимулов; фактически, значимая синхронизация проявилась лишь для комплексного стимула (t = 3,4; p < 0,001), а для элементных стимулов уровень синхронизации не отличается от фонового уровня (p > 0,05).
На графике динамики мощности тета-осцилляций (рис. 4B) видно, что мощность тета-осцилляций при комплексном подкрепляемом стимуле выше, чем у комплексного неподкрепляемого стимула, практически на всем анализируемом интервале.
Для более детального анализа значимые сенсоры были сгруппированы в пять пространственных топографических кластеров (рис. 5). Как видно на рисунке, общий характер выявленных закономерностей присутствует во всех выделенных областях. Реакция на подкрепляемый комплексный стимул статистически значимо выше, чем на неподкрепляемый, во всех областях, за исключением левой теменно-височной области. Комплексный стимул демонстрирует статистически значимо более высокую мощность тета-осцилляций по сравнению с элементными стимулами в условии подкрепления во всех выделенных пространственных кластерах.
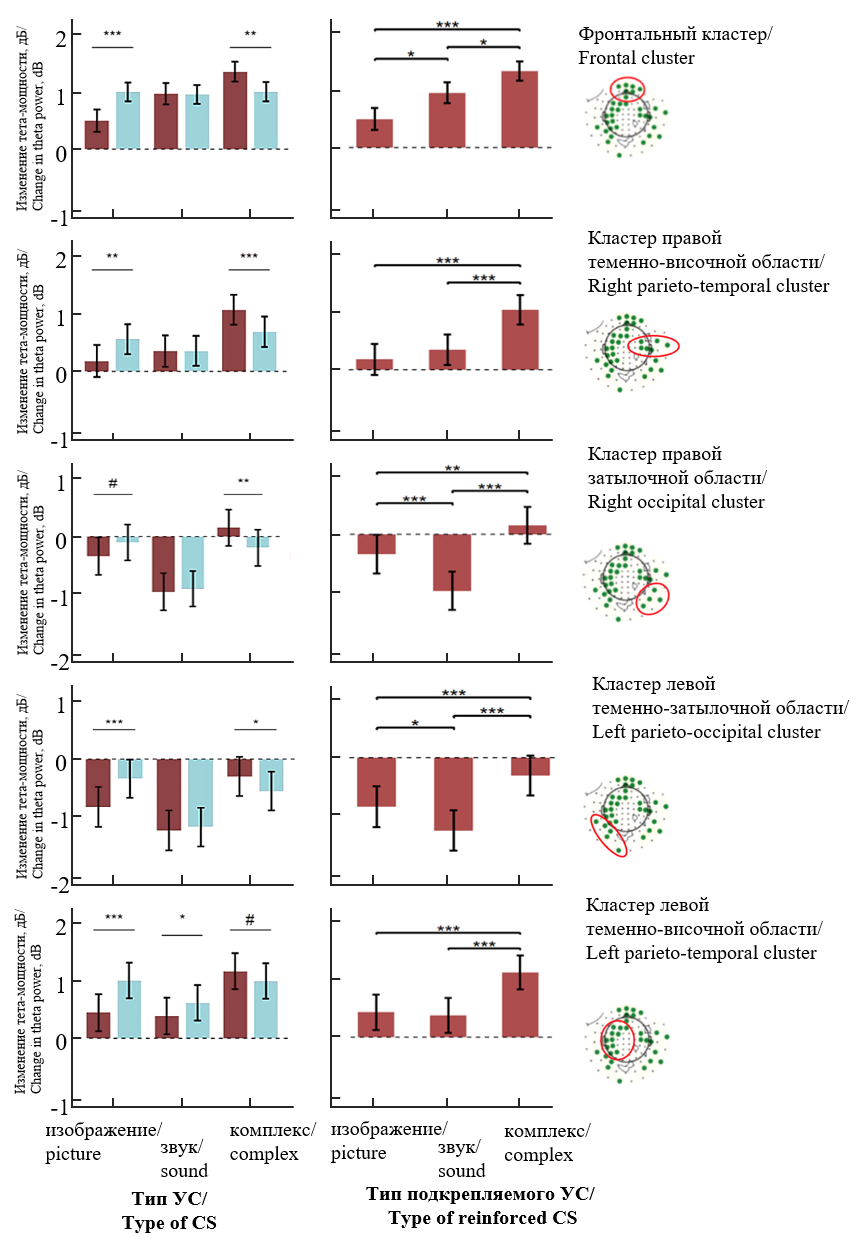
Рис. 5. Анализ мощности тета-осцилляций в кластерах сенсоров (достоверные сенсоры, выделенные зеленым цветом, обведены красным овалом на топографических развертках пространства сенсоров). Обозначения как на рис. 4
Fig. 5. Analysis of theta oscillation power in sensor clusters (significant sensors highlighted in green are outlined with a red oval on the topographic layouts of sensor space). Notations are the same as in Fig. 4
Обсуждение результатов
Участники эксперимента успешно выполняли экспериментальную задачу, включавшую выработку ассоциации как на конфигурации стимулов, так и на отдельные элементы этих конфигураций. Благодаря сбалансированному дизайну мы получили возможность сравнивать активность мозга в зависимости от типа стимула — элементного или комплексного, а также в зависимости от того, сочетался ли стимул с подкреплением или не сочетался.
Мы выявили зоны, в которых проявились два сопряженных эффекта: 1) мощность тета-осцилляций была выше в ответ на подкрепляемый комплексный стимул в сравнении с подкрепляемыми элементными стимулами; 2) мощность тета-осцилляций была выше в ответ на подкрепляемый комплексный стимул в сравнении с неподкрепляемым комплексным стимулом. Применение сбалансированного дизайна с четырьмя контрабалансировочными группами позволило нам снизить влияние потенциально возможных эффектов, связанных с физическими характеристиками стимула.
Важно отметить, что выявленные эффекты в отношении комплексных стимулов не могут быть объяснены простой суммацией вызванных полей на слуховой и зрительный компоненты комплексного стимула. Во-первых, данный факт подтверждается результатами примененного нами статистического подхода, который основан на поиске значимых кластеров по взаимодействию между факторами, характеризующими условный стимул и подкрепление, а не по соответствующим факторам в отдельности. Во-вторых, эффекты преобладания подкрепляемого комплексного стимула проявились не только в раннем интервале (в период генерации N1-P2), но и в более позднем интервале (рис. 4В), в котором регистрируются наиболее длиннолатентные вызванные поля (такие как P300), не связанные с физическими показателями стимуляции и отражающие когнитивные факторы (Polich, 2007).
Эффекты кодирования комплексного стимула, ассоциируемого с подкреплением, выявлены в нескольких локализациях, включая префронтальную кору билатерально (вероятно, преимущественно медиальную префронтальную), левые дорсолатеральные фронтальные области, правые височные области и теменно-затылочные области. Топография выявленных эффектов указывает на вовлечение высокоуровневых ассоциативных зон, включая префронтальные и заднетеменные, что соответствует результатам других исследований механизмов вовлечения данных ассоциативных зон в конфигурационное научение, проведенных с применением функциональной магниторезонансной томографии (Baeuchl et al., 2015; Sehlmeyer et al., 2009), электро- и магнитоэнцефалографии (Cashdollar et al., 2009; Fuentemilla et al., 2010; Olsen et al., 2013; Poch et al., 2011).
Полученные нами результаты подтверждают имеющееся в литературе мнение, что у человека кодирование ассоциаций на комплексные стимулы вовлекает осцилляции в тета-диапазоне (Cashdollar et al., 2009; Fuentemilla et al., 2010; Olsen et al., 2013; Poch et al., 2011). Однако в настоящей работе данный феномен впервые выявлен для межмодальных комплексных стимулов, поскольку во всех известных нам исследованиях на человеке применяли лишь внутримодальные комплексы (сложные зрительные стимулы).
В соответствии с выводами других научных работ в данной области (Jensen, Hennequin, Mattar, 2024; Joensen et al., 2023; Karakaş, 2020; Miller, 2013; Nardin et al., 2023), мы можем предположить, что наблюдаемое нами усиление тета-осцилляций отражает функции гиппокампа по связыванию элементов комплексного стимула — особенно взаимодействие между гиппокампом и ассоциативными областями коры в процессе укрепления и реализации такой ассоциации (Karakaş, 2020; Miller, 2013).
Заключение
Таким образом, в данном исследовании впервые показано, что кодирование конфигурационной межмодальной ассоциации сопряжено с наибольшей выраженностью тета-осцилляций. Предположительно, этот феномен является отражением вовлечения гиппокампа в кодирование комплексного стимула и его взаимодействия c ассоциативными зонами коры.
Литература
- Ивашкина, О.И., Торопова, К.А., Рощина, М.А., Анохин, К.В. (2020). Формирование и извлечение ассоциативной памяти на комплексный сигнал у мышей: специфическое участие нейронов области СА1 гиппокампа. Журнал высшей нервной деятельности им. И.П. Павлова, 70(3), 327—341.
Ivashkina, O.I., Toropova, K.A., Roshchina, M.A., Anokhin, K.V. (2020). Formirovanie i izvlechenie assotsiativnoi pamyati na kompleksnyi signal u myshei: spetsificheskoe uchastie neironov oblasti SA1 gippokampa. Zhurnal vysshei nervnoi deyatel'nosti im. I.P. Pavlova, 70(3), 327—341. - Чернышев, Б.В., Ушаков, В.Л., Позняк, Л.А. (2024). Поиск нейрофизиологических механизмов конфигурационного обучения. Журнал высшей нервной деятельности им. И.П. Павлова, 74(2), 149—165.
Chernyshev, B.V., Ushakov, V.L., Poznyak, L.A. (2024). Poisk neirofiziologicheskikh mekhanizmov konfiguratsionnogo obucheniya. Zhurnal vysshei nervnoi deyatel'nosti im. I.P. Pavlova, 74(2), 149—165. - Baeuchl, C., Meyer, P., Hoppstädter, M., Diener, C., Flor, H. (2015). Contextual fear conditioning in humans using feature-identical contexts. Neurobiology of Learning and Memory, 121, 1—11. https://doi.org/1016/j.nlm.2015.03.001
- Bronfman, Z.Z., Ginsburg, S., Jablonka, E.М. (2016). The Transition to Minimal Consciousness through the Evolution of Associative Learning. Frontiers in Psychology, 7. https://doi.org/3389/fpsyg.2016.01954
- Buzsáki, G., McKenzie, S., Davachi, L. (2022). Neurophysiology of Remembering. Annual Review of Psychology, 73, 187—215. https://doi.org/1146/annurev-psych-021721-110002
- Cashdollar, N., Malecki, U., Rugg-Gunn, F.J., Duncan, J.S., Lavie, N., Duzel, E. (2009). Hippocampus-dependent and -independent theta-networks of active maintenance. Proceedings of the National Academy of Sciences, 106(48), 20493—20498. https://doi.org/1073/pnas.0904823106
- Duncan, K., Doll, B.B., Daw, N.D., Shohamy, D. (2018). More Than the Sum of Its Parts: A Role for the Hippocampus in Configural Reinforcement Learning. Neuron, 98(3), 645—657. https://doi.org/10.1016/j.neuron.2018.03.042
- Feinberg, T.E., Mallatt, J. (2016). The nature of primary consciousness. A new synthesis. Consciousness and Cognition, 43, 113—127. https://doi.org/10.1016/j.concog.2016.05.009
- Fuentemilla, L., Penny, W.D., Cashdollar, N., Bunzeck, N., Düzel, E. (2010). Theta-Coupled Periodic Replay in Working Memory. Current Biology, 20(7), 606—612. https://doi.org/10.1016/j.cub.2010.01.057
- Ginsburg, S., Jablonka, E. (2019). The evolution of the sensitive soul: learning and the origins of consciousness. Cambridge, MA: MIT Press.
- Gramfort, A., Luessi, M., Larson, E., Engemann, D.A., Strohmeier, D., Brodbeck, C., Parkkonen, L., Hämäläinen, M.S. (2014). MNE software for processing MEG and EEG data. NeuroImage, 86, 446—460. https://doi.org/10.1016/j.neuroimage.2013.10.027
- Jensen, K.T., Hennequin, G., Mattar, M.G. (2024). A recurrent network model of planning explains hippocampal replay and human behavior. Nature Neuroscience. https://doi.org/10.1038/s41593-024-01675-7
- Joensen, B.H., Bush, D., Vivekananda, U., Horner, A.J., Bisby, J.A., Diehl, B., Miserocchi, A., McEvoy, A.W., Walker, M.C., Burgess, N. (2023). Hippocampal theta activity during encoding promotes subsequent associative memory in humans. Cerebral Cortex, 33(13), 8792—8802. https://doi.org/10.1093/cercor/bhad162
- Karakaş, S. (2020). A review of theta oscillation and its functional correlates. International Journal of Psychophysiology. https://doi.org/10.1016/j.ijpsycho.2020.04.008
- Kimchi, R. (1994). The Role of Wholistic/Configural Properties versus Global Properties in Visual Form Perception. Perception, 23(5), 489—504. https://doi.org/10.1068/p230489
- Lisman, J., Buzsaki, G. (2008). A neural coding scheme formed by the combined function of gamma and theta oscillations. Bull., 34(5), 974—980.
- Luck, S.J. (2014). An introduction to the event-related potential technique. Cambridge, MA: MIT press.
- Maren, S., Aharonov, G., Fanselow, M.S. (1997). Neurotoxic lesions of the dorsal hippocampus and Pavlovian fear conditioning in rats. Behavioural Brain Research, 88(2), 261—274. https://doi.org/10.1016/S0166-4328(97)00088-0
- Maren, S., Phan, K.L., Liberzon, I. (2013). The contextual brain: implications for fear conditioning, extinction and psychopathology. Nature Reviews Neuroscience, 14(6), 417—428. https://doi.org/10.1038/nrn3492
- Miller, R. (2013). Cortico-hippocampal interplay and the representation of contexts in the brain. Springer Science & Business Media.
- Nardin, M., Kaefer, K., Stella, F., Csicsvari, J. (2023). Theta oscillations as a substrate for medial prefrontal-hippocampal assembly interactions. Cell Reports, 42(9), 113015. https://doi.org/10.1016/j.celre 2023.113015
- Navon, D. (1977). Forest before trees: The precedence of global features in visual perception. Cognitive Psychology, 9(3), 353—383. https://doi.org/10.1016/0010-0285(77)90012-3
- Olsen, R.K., Rondina, Ii R., Riggs, L., Meltzer, J.A., Ryan, J.D. (2013). Hippocampal and neocortical oscillatory contributions to visuospatial binding and comparison. Journal of Experimental Psychology: General, 142(4), 1335—1345. https://doi.org/10.1037/a0034043
- Poch, C., Fuentemilla, L., Barnes, G.R., Düzel, E. (2011). Hippocampal Theta-Phase Modulation of Replay Correlates with Configural-Relational Short-Term Memory Performance. The Journal of Neuroscience, 31(19), 7038—7042. https://doi.org/10.1523/jneurosci.6305-10.2011.
- Polich, J. (2007). Updating P300: An integrative theory of P3a and P3b. Clinical Neurophysiology, 118(10), 2128—2148. https://doi.org/10.1016/j.clinph.2007.04.019
- Razorenova, A.M., Chernyshev, B.V., Nikolaeva, A.Y., Butorina, A.V., Prokofyev, A.O., Tyulenev, N.B., Stroganova, T.A. (2020). Rapid Cortical Plasticity Induced by Active Associative Learning of Novel Words in Human Adults. Frontiers in Neuroscience, 14, https://doi.org/10.3389/fnins.2020.00895
- Razran, G. (1971). Mind in evolution: An East-West synthesis of learned behavior and cognition. Houghton Mifflin.
- Rudy, J.W., Huff, N.C., Matus-Amat P. (2004). Understanding contextual fear conditioning: insights from a two-process model. Neuroscience & Biobehavioral Reviews, 28(7), 675—685. https://doi.org/10.1016/j.neubiorev.2004.09.004
- Rudy, J.W., Sutherland, R.J. (1995). Configural association theory and the hippocampal formation: An appraisal and reconfiguration. Hippocampus, 5(5), 375—389. https://doi.org/10.1002/hipo.450050502
- Sakimoto, Y., Hattori, M., Takeda, K., Okada, K., Sakata, S. (2013). Hippocampal theta wave activity during configural and non-configural tasks in rats. Experimental Brain Research, 225(2), 177—185. https://doi.org/10.1007/s00221-012-3359-2
- Sakimoto, Y., Okada, K., Takeda, K., Sakata, S. ( 2013). Transient Decline in Hippocampal Theta Activity during the Acquisition Process of the Negative Patterning Task. PLOS ONE, 8(7), e70756. https://doi.org/10.1371/journal.pone.0070756
- Sakimoto, Y., Sakata, S. (2015). The transient decline in hippocampal theta power during response inhibition in a positive patterning task. NeuroReport, 26(14), 833—837. https://doi.org/10.1097/wnr.0000000000000432
- Sehlmeyer, C., Schöning, S., Zwitserlood, P., Pfleiderer, B., Kircher, T., Arolt, V., Konrad, C. (2009). Human Fear Conditioning and Extinction in Neuroimaging: A Systematic Review. PLOS ONE, 4(6), https://doi.org/10.1371/journal.pone.0005865
- Stout, D.M., Glenn, D.E., Acheson, D.T., Simmons, A.N., Risbrough, V.B. (2019). Characterizing the neural circuitry associated with configural threat learning. Brain Research, 1719, 225—234. https://doi.org/10.1016/j.brainres.2019.06.003
- Stout, D.M., Glenn, D.E., Acheson, D.T., Spadoni, A.D., Risbrough, V.B., Simmons, A.N. (2018). Neural measures associated with configural threat acquisition. Neurobiology of Learning and Memory, 150, 99—106. https://doi.org/10.1016/j.nlm.2018.03.012
- Sutherland, R.J., Rudy, J.W. (1989). Configural association theory: The role of the hippocampal formation in learning, memory, and amnesia. Psychobiology, 17(2), 129—144. https://doi.org/10.3758/BF03337828
Информация об авторах
Метрики
Просмотров web
За все время: 15
В прошлом месяце: 0
В текущем месяце: 15
Скачиваний PDF
За все время: 2
В прошлом месяце: 0
В текущем месяце: 2
Всего
За все время: 17
В прошлом месяце: 0
В текущем месяце: 17