Экспериментальная психология
2014. Том 7. № 3. С. 31–43
ISSN: 2072-7593 / 2311-7036 (online)
Научение и поведение в отсутствие зрительного контакта со средой у крыс*
Аннотация
Общая информация
* Работа выполнена при поддержке Российского научного фонда (проект №14-08-00229 «Системная дифференциация как основа биологического и социокультурного развития»).
Ключевые слова: индивидуальный опыт, научение, поведение, зрение, крыса
Рубрика издания: Когнитивная психология
Тип материала: научная статья
Для цитаты: Арутюнова К.Р., Гаврилов В.В., Александров Ю.И. Научение и поведение в отсутствие зрительного контакта со средой у крыс // Экспериментальная психология. 2014. Том 7. № 3. С. 31–43.
Полный текст
Введение
Зрению придается исключительное значение в организации отношения индивида со средой (Барабанщиков, Жегалло, 2013). Традиционно под зрением понимаются механизмы переработки зрительной информации о среде в поведении (Marr, 1982; Riesenhuber, Poggio, 2002; Ungerleider, Mishkin, 1982 и др.), однако такой подход в современных исследованиях подвергается критике в силу его ограниченности и при попытках описания общих механизмов взаимодействия индивида с миром (см. обзор в: Cisek, Kalaska, 2010). С позиций системноэволюционного подхода в психофизиологии (Александров, 1989; Александров и др., 1997; Швырков, 1978, 1986, 1995), мы рассматриваем зрение как сформировавшуюся в эволюции возможность использовать определенные оптические параметры среды для достижения адаптивных поведенческих результатов. Зрение здесь рассматривается не как функция, а как характеристика целостного поведения организма в аспекте его соотношения со «зрительными» свойствами среды. При этом то же самое поведение в зависимости от целей исследования может рассматриваться с другой стороны – в соотношении со «слуховыми», «обонятельными» и другими свойствами этой среды. При нормальном развитии и соответствующей видовой обусловленности зрение является неотъемлемой частью формирования индивидуального опыта – приобретения и фиксации адаптивных моделей взаимодействия индивида с миром. В данной работе изучалось, как наличие и отсутствие контакта с оптическими параметрами среды может проявляться в формировании и реализации индивидуального опыта у крыс при научении новому поведению и его реализации в разных условиях.
Несмотря на то, что, ввиду ночного образа жизни и особенностей естественной среды обитания, экологическое значение зрения для крыс не настолько высоко, как, например, обоняния и слуха, эти животные обладают типичной для млекопитающих зрительной системой и успешно обучаются решению целого ряда сложных зрительных задач (Prusky, Douglas, 2005), в том числе и аналогичных тем, которые используют в исследованиях на приматах и человеке (Prusky et al., 2004; Vermaercke, Op de Beeck, 2012). Показано, что у грызунов скорость обучения некоторым задачам связана с остротой зрения, которая в целом играет значимую роль в пространственном обучении и памяти (Brown, Wong, 2007). При этом известно, что пигментированные крысы, в частности линия Long Evans, обладают более острым зрением, чем альбиносные формы лабораторных крыс (Prusky et al., 2002), поэтому первые способны использовать его более эффективно при решении ряда поведенческих задач (Tonkiss et al., 1992). В данной работе применялась модель инструментального поведения, в которой животные, с одной стороны, могли обучиться без зрения, а с другой – имели возможность использовать зрительные ориентиры, если их зрение не ограничивалось условиями эксперимента.
Изучение того, как отсутствие зрительного контакта со средой сказывается на поведении и его мозговой организации, часто связано с исследованием структурных и функциональных изменений при слепоте и долговременной зрительной депривации у животных (см., напр.: Bavelier, Neville, 2002; Hill, Best, 1981; Save et al., 1998; Zernicki, 1979 и др.) и человека (Сергиенко, 1995; Amedi et al., 2003; Bavelier, Neville, 2002; Lambert et al., 2004 и др.) Целью данного исследования, напротив, было выявление поведенческих особенностей научения при исключении зрительного контакта со средой у крыс с интактной зрительной системой и нормальным зрительным развитием. Для этого изучались поведенческие показатели научения животных новой задаче в незнакомой среде с открытыми и с закрытыми глазами, а также его последующая реализация в этих и обратных условиях. Исходя из системно-эволюционных представлений, мы полагаем, что зрительный контакт со средой может быть связан с обеспечением большей степени дифференцированности характеристик структуры индивидуального опыта, тем самым обеспечивая более сложные способы взаимодействия индивида со средой. Исключение возможности использовать оптические параметры среды на поведенческом уровне может проявляться в скорости и динамике научения, а также при реализации актов уже сформированного поведения. Для выявления и анализа возможных особенностей научения и поведения в отсутствие зрительного контакта со средой у крыс в данной работе сопоставляются количественные показатели научения и поведения в двух группах животных, обучающихся одному и тому же инструментальному поведению в разных условиях – с открытыми и с закрытыми глазами.
Методика
Исследование проведено на 47 крысах линии Long Evans в возрасте 3–11 месяцев. Крысы содержались в отдельных клетках в вивариуме с 12-часовым циклом смены дня и ночи. Для обучения инструментальному пищедобывательному поведению все крысы были разделены на две группы. Животные первой группы (n=31) попадали в комнату, где проводился эксперимент, с уже закрытыми глазами, и до этого они никогда эту комнату не видели. Глаза закрывались специально сконструированными светонепроницаемыми колпачками (рис. 1А). Колпачки надевали лишь на 30–40 мин в день – только на время проведения экспериментальных сессий, в домашних клетках животные находились с открытыми глазами. Животные второй группы (n=16) обучались с открытыми глазами. Перед каждой сессией с животными этой группы проводились манипуляции по надеванию колпачков, аналогичные проводившимся с животными первой группы.
Для контроля качества закрытия глаз проверяли наличие вызванных потенциалов (ВП) на ритмические вспышки света в корковых отведениях ЭЭГ (рис. 1Б). Для этого всем животным до начала обучения под общей анестезией (Zoletil, внутримышечно) были имплантированы хлорсеребряные электроды (сопротивление – 30 кОм). Электроды располагались эпидурально над моторной (а. 3.0, l. 3.5), ретросплениальной (p. 4.5, l. 1.0) и зрительной (p. 7.5, l. 4.0) областями коры правого полушария мозга. Референтный электрод располагался над носовыми пазухами (а. 11.0, l. 1.0), землей служил электрод, расположенный под затылочной костью. Эксперимент начинали через 7–10 дней после операции, что требовалось для восстановления животных. Генерация вспышек света при регистрации ВП производилась при помощи бесшумной светодиодной лампы, расположенной на потолке клетки, на расстоянии 30–50 см от животного. Вспышки предъявлялись в течение двух мин, при этом крыса находилась в свободном поведении в боксе размером 30 x 40 см на расстоянии 50 см от источника вспышек. Частота вспышек – 1 Гц, длительность – 30 мс. Для усиления сигнала ЭЭГ использовали четырехканальный усилитель DL304N NBLab. Полоса пропускания – 1 Гц–1 КГц.
Исследование проводилось в экспериментальной клетке, которая представляла собой куб с ребром 60 см. По углам клетки располагались две педали и две автоматически подающиеся кормушки. При нажатии на эффективную педаль порция пищи (кубик сыра) подавалась в соответствующую данной педали кормушку. Эффективной могла быть только одна педаль. Смена эффективной педали производилась экспериментатором. Экспериментальная клетка была оборудована датчиками, позволяющими регистрировать моменты и продолжительность нажатия на педали и опускания головы животного в кормушки.
В экспериментальной клетке животные имели возможность научиться нажимать на педаль для получения пищи в кормушке. До начала сессий обучения животные подвергались пищевой депривации (потеря в весе не превышала 20% от первоначального веса животного). Методом проб и ошибок крысы научались совершать инструментальное пищедобывательное поведение сначала на одной педали, затем на второй. Для этого крысу помещали в экспериментальную клетку на 30 мин ежедневно. Критерием научения служило регулярное циклическое поведение нажатия на педаль и получения пищи в кормушке. Объективным показателем научения служило число нажатий на эффективную педаль с последующими подходами к кормушке. Считали, что животное научилось, когда это число превышало 30 в одну сессию обучения, и поведение становилось цикличным: нажатие на педаль – побежка к кормушке – залезание в кормушку и захват пищи – побежка к педали и т.д. Когда животное научалось поведению на двух педалях клетки, в отдельных экспериментальных сессиях проводили регистрацию поведенческих показателей.
Поведенческие показатели проанализированы у 18 животных (по 9 животных из каждой группы). У этих животных регистрацию поведения проводили в три этапа. На первом этапе поведение тестировалось в тех же условиях, в которых оно формировалось, с закрытыми или с открытыми глазами. На втором этапе животным, которые обучались с закрытыми глазами, светонепроницаемые колпачки снимали, а животным, которые обучались с открытыми глазами, – колпачки надевали. После восстановления регулярного циклического поведения в новых для животных условиях проводили регистрацию поведенческих показателей. Третьим этапом считали повторное надевание колпачков животным первой группы, которые обучались с закрытыми глазами, и повторное снятие колпачков животным второй группы, которые обучались с открытыми глазами. Регистрировали поведение в тех же условиях, в которых животные обучались изначально.
Для анализа данных использовали программы IBM SPSS Statistics 20 и Statistica 6.0. При сопоставлении скорости научения и времени реализации актов поведения между животными двух групп использовали U-критерий Манна-Уитни. Для анализа времени реализации поведения на разных этапах эксперимента считали медианное значение скорости актов поведения и разброс в квартилях. Сдвиг в показателе времени реализации актов поведения на разных этапах эксперимента оценивали по критерию Т Вилкоксона. Достоверными считали различия с уровнем значимости p<0,05.
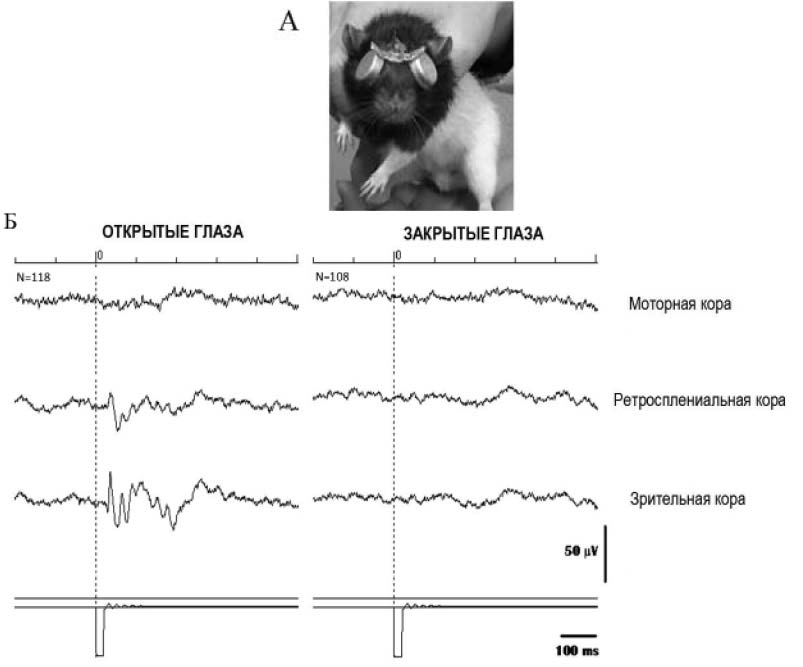
Рис. 1. Исключение зрительного контакта со средой. А. Крыса с надетыми на глаза светонепроницаемыми колпачками. Б. ЭЭГ-потенциалы на ритмические вспышки света. Показаны усредненные потенциалы из отведений в моторной, ретросплениальной и зрительной коре крысы с открытыми (слева) и закрытыми (справа) глаза ми. С открытыми глазами наблюдались вызванные потенциалы (ВП) в зрительной и ретросплениальной коре. С закрытыми глазами ВП отсутствовали
Результаты
Большинство животных (68%, n=21 – в группе с закрытыми глазами и 81%, n=13 – в группе с открытыми глазами, см. верхние диаграммы на рис. 2) научились нажимать на педали для получения пищи в кормушках. Число животных, не обучившихся данному поведению за отведённые 15 тридцатиминутных сессий, в двух группах значимо не отличалось (см. верхние диаграммы на рис. 2: χ2(1, N=47)=0,96, p=0,33).
Для обучения на первой педали животным обеих групп потребовалось от 4-х до 14-и тридцатиминутных сессий, на второй – от 1-й до 4-х. Все животные обучались поведению на второй педали быстрее, чем на первой, продемонстрировав «перенос» навыка. Разницы между двумя группами в скорости научения не было обнаружено ни на первой (критерий Манна-Уитни, n1=21, n2=13, U=123,5, p=0,65), ни на второй (критерий Манна-Уитни, n1=21, n2=13, U=74,5, p=0,09) педалях (см. нижние гистограммы на рис. 2).
Хотя животным потребовалось разное количество сессий для освоения задачи на первой стороне экспериментальной клетки, динамика научения была сходной у крыс двух групп (рис. 3). В первую сессию обучения, как правило, наблюдалось активное исследовательское поведение, в процессе которого животные многократно совершали побежки по периметру клетки, несколько раз нажимая на эффективную педаль и затем находя пищу в соответствующей данной педали кормушке. В последующие сессии обучения количество нажатий на педали и активность животных несколько снижались. Значимое повышение эффективности поведения, соответствующее использованному нами объективному критерию научения (см. Методика), как правило, происходило в рамках одной сессии и наступало на 4–14-й день. С животными, не обучившимися за 15 тридцатиминутных сессий, эксперимент прекращали. Зачастую их поведение можно было охарактеризовать как «пассивное ожидание» окончания сессии и возвращения в домашние клетки, их активность была значительно снижена и сводилась к нахождению в одном из углов экспериментальной клетки и грумингу. Тем не менее, это не исключает возможности того, что и эти животные могли бы научиться задаче, если бы им дали возможность находиться в экспериментальной клетке в течение большего количества сессий.
Обучение на второй стороне было значительно динамичнее. После некоторого количества неуспешных попыток нажатия на неэффективную педаль, животные совершали побежки по периметру клетки и довольно быстро находили вторую педаль, которая становилась эффективной и приносила пищу в другой кормушке. При этом, несмотря на быстрое обнаружение эффективности второй педали, животные часто возвращались к первой педали и совершали по нескольку проб на неэффективной стороне.
Научившиеся задаче животные обеих групп реализовывали сходное по структуре циклическое поведение, в котором для последующего анализа выделялись следующие акты: нажатие на педаль, побежка к кормушке, поедание пищи в кормушке и побежка к педали. При анализе длительностей актов нажатия на педаль и поедания пищи в кормушке наблюдалась высокая вариативность как между реализациями данных актов разными животными, так и в отдельных циклах, реализуемых одними и теми же животными. Принимая во внимание данную вариативность, для выявления особенностей в скорости реализации поведения с использованием зрения и при его исключении сопоставлялись только длительности побежек от педалей к кормушкам и от кормушек к педалям.
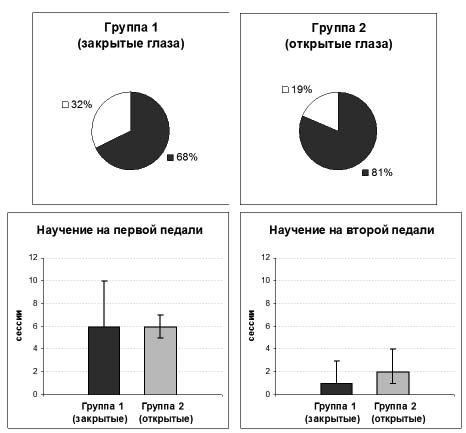
Рис. 2. Научение инструментальному поведению у двух групп животных. На верхних диаграммах отражено процентное соотношение числа животных, научившихся (темно-серым цветом) и не научившихся задаче. Слева – группа, обучавшая с закрытыми глазами (n=31), справа – с открытыми глазами (n=16). Соотношение числа научившихся и не научившихся животных между двумя группами значимо не отличалось (критерий χ2, p>0,05). На нижних гистограммах указано число тридцатиминутных сессий, потребовавшееся для научения животных поведению на первой (слева) и второй (справа) педалях. Указаны медианные значения и разброс в квартилях. Группа 1 (n=21) обозначена темно-серым цветом, группа 2 (n=13) – светло-серым цветом. Значимых различий в скорости научения между двумя группами обнаружено не было (критерий Манна-Уитни, p>0,05)
Анализ поведенческих показателей в дефинитивном поведении после научения задаче показал, что животные с открытыми глазами (группа 2) совершали побежки к кормушкам и к педалям быстрее, чем животные с закрытыми глазами (группа 1), это наблюдалось как на первой (критерий Манна-Уитни, n1=9, n2=9, U=18, p<0,05), так и на второй (критерий Манна-Уитни, n1=9, n2=9, U=16, p<0,03) сторонах экспериментальной клетки (рис. 4). У животных обеих групп скорость реализации поведения на первой и второй сторонах клетки не отличалась (критерий Вилкоксона, p>0,05).
При анализе поведения на трех этапах эксперимента у животных, обучавшихся с открытыми и с закрытыми глазами, наблюдалась разная динамика. У крыс из группы 2, изначально обучавшихся с открытыми глазами, закрывание глаз на втором этапе эксперимента приводило к значимому замедлению поведения (рис. 5, критерий Вилкоксона, первая сторона: n=9, Z=2,55, p<0,02; вторая сторона: n=9, Z=2,67, p<0,01). При этом, после открывания глаз на третьем этапе скорость реализации побежек снова увеличивалась до прежнего уровня (критерий Вилкоксона, первая сторона: n=9, Z=2,55, p<0,02; вторая сторона: n=9, Z=2,67, p<0,01). В группе 1 наблюдалась менее выраженная динамика скорости реализации поведения на протяжении трех этапов эксперимента. У животных, обучавшихся с закрытыми глазами, открывание глаз на втором этапе эксперимента привело к небольшому ускорению поведения, и этот эффект достиг уровня значимости только на первой стороне экспериментальной клетки (критерий Вилкоксона, n=9, Z=2,31, p<0,03). Повторное закрывание глаз не приводило к значимым сдвигам в скорости поведения по сравнению с первым и вторым этапами (критерий Вилкоксона, p>0,05).
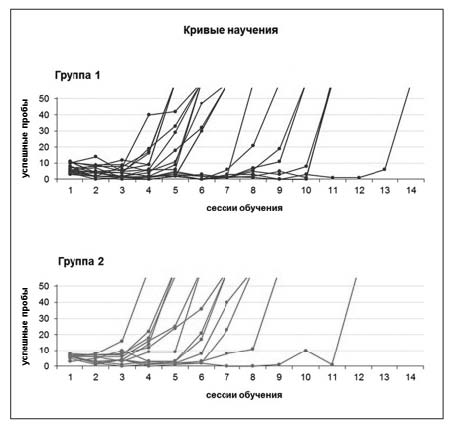
Рис. 3. Кривые научения задаче для животных двух групп. Показано число успешных проб – нажатий на эффективную педаль с последующим захватом пищи – в сессиях научения на первой стороне экспериментальной клетки. Животным потребовалось разное количество сессий для освоения задачи – от 4-х до 14-и, однако динамика научения была сходной у крыс двух групп: значимое повышение эффективности поведения, как правило, происходило в рамках одной сессии. Объективный показатель научения – число нажатий на эффективную педаль с последующими подходами к кормушке. Считали, что животное научилось, когда это число превышало 30 в одну сессию обучения, и поведение становилось цикличным
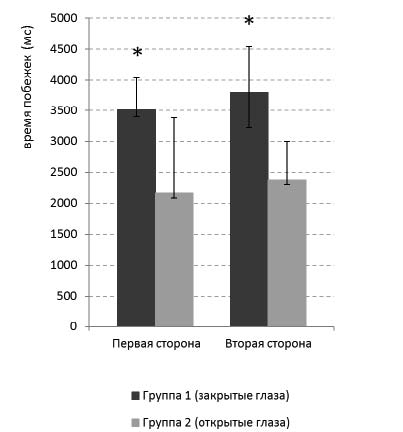
Рис. 4. Скорость реализации поведения после научения. Время побежек к педалям и кормушкам клетки
(в мс) после обучения на обеих сторонах экспериментальной клетки. Темно-серые столбцы – поведение животных с закрытыми глазами (группа 1). Светло-серые столбцы – с открытыми глазами (группа 2). На обеих сторонах клетки животные с открытыми глазами совершают поведение быстрее, чем животные с закрытыми глазами, критерий Манна-Уитни, *p<0,05
При анализе поведения на трех этапах эксперимента у животных, обучавшихся с открытыми и с закрытыми глазами, наблюдалась разная динамика. У крыс из группы 2, изначально обучавшихся с открытыми глазами, закрывание глаз на втором этапе эксперимента приводило к значимому замедлению поведения (рис. 5, критерий Вилкоксона, первая сторона: n=9, Z=2,55, p<0,02; вторая сторона: n=9, Z=2,67, p<0,01). При этом, после открывания глаз на третьем этапе скорость реализации побежек снова увеличивалась до прежнего уровня (критерий Вилкоксона, первая сторона: n=9, Z=2,55, p<0,02; вторая сторона: n=9, Z=2,67, p<0,01). В группе 1 наблюдалась менее выраженная динамика скорости реализации поведения на протяжении трех этапов эксперимента. У животных, обучавшихся с закрытыми глазами, открывание глаз на втором этапе эксперимента привело к небольшому ускорению поведения, и этот эффект достиг уровня значимости только на первой стороне экспериментальной клетки (критерий Вилкоксона, n=9, Z=2,31, p<0,03). Повторное закрывание глаз не приводило к значимым сдвигам в скорости поведения по сравнению с первым и вторым этапами (критерий Вилкоксона, p>0,05).
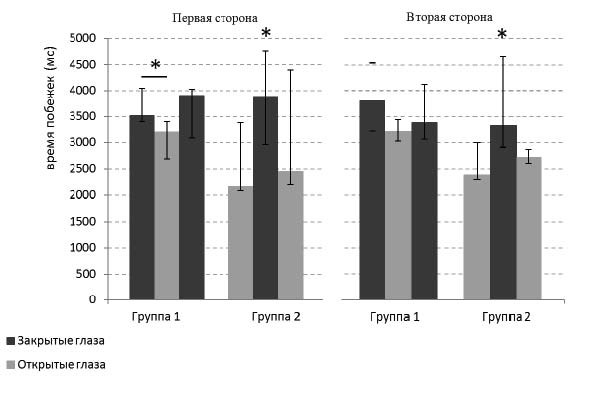
Рис. 5. Динамика реализации поведения на трех стадиях эксперимента. Гистограммы времени побежек от педалей к кормушкам и обратно для двух групп животных на трех стадиях эксперимента, показаны медианные значения и разброс в квартилях. Темно-серые столбцы – закрытые глаза, светло-серые – открытые. Критерий Манна-Уитни, различия при *p<0,05
Обсуждение результатов
Полученные результаты показали, что исключение зрительного контакта со средой не влияет значимо на успешность научения крыс новому инструментальному поведению в незнакомой среде. По скорости и динамике научения животные с закрытыми глазами не отличались от животных, обучавшихся с открытыми глазами. В условиях отсутствия возможности использовать оптические параметры среды крысы могут эффективно решать поведенческие задачи через другие способы взаимодействия со средой – осязание, обоняние, слух и пр. (Prusky, Douglas, 2005). Кроме того, показано, что в темноте топографическая организация исследовательского поведения крыс не отличается от таковой на свету (Whishaw et al., 2001). В используемой нами задаче при научении животных без помощи экспериментатора исследовательское поведение, особенно в первые сессии нахождения в новой экспериментальной среде, играет основополагающую роль для последующего формирования навыка нажатия на педали для получения пищи в кормушках. Поэтому сходство исследовательских стратегий данного вида вне зависимости от возможности использовать оптические параметры среды, а также присутствие множества других средовых ориентиров – слуховых (звуки кормушек и педалей), обонятельных (запах кормушек и собственных меток), проприоцептивных (ощущения собственных движений в пространстве экспериментальной клетки) и др. – может обусловливать отсутствие значимых различий в скорости и динамике научения крыс инструментальному поведению.
Однако, при общем сходстве процесса научения задаче, были показаны некоторые различия при реализации актов уже сформированного поведения: на первом этапе животные с закрытыми глазами выполняли побежки к педалям и кормушкам медленнее, чем животные с открытыми глазами, а затем, на последующих этапах эксперимента при смене условий наблюдалась разная динамика скорости реализации актов у животных, обучавшихся одному и тому же поведению с закрытыми и с открытыми глазами. С одной стороны, полученные данные согласуются с литературой о том, что скорость движений и перемещений крыс в темноте в целом ниже, чем на свету (Whishaw et al., 2001). С другой стороны, показано, что у животных в одной и той же экспериментальной среде при совершении одного и того же по структуре поведения в двух разных условиях – при неограниченной возможности использовать зрение и при его исключении – у части нейронов наблюдаются схожие активации, а у других нейронов – значимые различия в паттернах активаций (Александров, Александров, 1981; Quirk et al., 1990) и др. Эти данные могут свидетельствовать о том, что одно и то же поведение с использованием зрения и без него обеспечивается перекрывающимися составами систем нейронов, где сходные активации связаны с актуализацией элементов опыта, обеспечивающих реализацию общей структуры поведения, а отличающиеся – с элементами опыта, лежащими в основе особенностей реализации актов поведения в специфических условиях, т.е. с использованием зрения или без него. Таким образом, особенности в системной организации индивидуального опыта, которая связана, в том числе, и с историей обучения данной задаче (с открытыми или с закрытыми глазами), также могут лежать в основе наблюдаемых различий в скорости и динамике реализации актов поведения.
Исходя из системно-эволюционных представлений, можно предположить, что даже в ситуации, когда оптические параметры среды не являются определяющими для обучения задаче и ее успешного выполнения, они, тем не менее, являются важным аспектом формирования и реализации индивидуального опыта. Мы полагаем, что в норме зрительный контакт со средой представляет собой одно из условий формирования любого нового опыта у организмов, обладающих соответствующими физиологическими структурами. При исключении данного условия формирование нового опыта будет происходить на основе уже имеющегося опыта, включающего «зрительный компонент». Однако этот новый опыт будет отличаться от всего предыдущего, и природа данного отличия может быть связана с особенностями отношений между элементами нового и старого опыта. Для изучения этих особенностей на системном уровне необходим анализ активности нейронов при формировании и реализации поведенческих актов в нормальных условиях и в отсутствие зрительного контакта со средой.
Литература
- Александров Ю.И. Психофизиологическое значение активности центральных и периферических нейронов в поведении. М.: Наука, 1989.
- Александров Ю.И., Александров И.О. Активность нейронов зрительной и моторной областей коры мозга при осуществлении поведенческого акта с открытыми и с закрытыми глазами //Журнал высшей нервной деятельности им. И.П. Павлова. 1981. Т. 31. № 6. С. 1179-1189.
- Александров Ю.И., Греченко Т.Н., Гаврилов В.В., Горкин А.Г., Шевченко Д.Г., Гринченко Ю.В., Александров И.О., Максимова Н.Е., Безденежных Б.Н., Бодунов М.В. Закономерности формирования и реализация индивидуального опыта // Журнал высшей нервной деятельности им. И.П. Павлова. 1997. Т. 47. № 2. С. 243-260.
- Барабанщиков В.А., Жегалло А.В. Регистрация и анализ направленности взора человека. М.: Изд-во Института Психологии РАН, 2013.
- Сергиенко Е.А. Влияние ранней зрительной депривации на интерсенсорное взаимодействие // Психологический журнал. 1995. Т. 16. № 5. С. 32-49.
- Швырков В.Б. Нейрофизиологическое изучение системных механизмов поведения. М.: Наука, 1978.
- Швырков В.Б. Изучение активности нейронов как метод психофизиологического исследования поведения // Нейроны в поведении: системные аспекты. М.: Наука, 1986. С. 6-25.
- Швырков В.Б. Введение в объективную психологию: нейрональные основы психики. М.: Изд-во Института Психологии РАН, 1995.
- Amedi A., Raz N., Pianka P., Malach R., Zohary E. Early 'visual' cortex activation correlates with superior verbal memory performance in the blind // Nature Neuroscience. 2003. V. 6. № 7. P. 758-776. doi:10.1038/nn1072.
- Bavelier D., Neville H.J. Cross-modal plasticity: where and how? // Nature Reviews Neuroscience. 2002. V. 3. P. 443-452. doi:10.1038/nrn848.
- Brown R.E., Wong A.A. The influence of visual ability on learning and memory performance in 13 strains of mice // Learning and Memory. 2007. V. 14. P. 134-144. doi: 10.1101/lm.473907.
- Cisek, P., Kalaska, J.F. Neural mechanisms for interacting with a world full of action choices // Annual Review of Neuroscience. 2010. V. 33. P. 269-298. doi: 10.1146/annurev.neuro.051508.135409.
- Hill A.J., Best P.J. Effects of deafness and blindness on the spatial correlates of hippocampal unit activity in the rat // Experimental Neurology. 1981. V. 74. P. 204-217. doi: 10.1016/0014-4886(81)90159-X.
- Lambert S., Sampaio E., Mauss Y., Scheiber C. Blindness and brain plasticity: contribution of mental imagery? An fMRI study // Cognitive Brain Research. 2004. V. 20. P. 1-11.
- Marr D.C. Vision: A computational investigation into the human representation and processing of visual information. New York: Freeman, 1982.
- Prusky G.T., Douglas R.M. Vision // The behaviour of the laboratory rat: a handbook with tests / Whishaw I.Q., Kolb B. (Eds.) New York: Oxford University Press, 2005. P. 49-59.
- Prusky G.T., Douglas R.M., Nelson L., Shabanpoor A., Sutherland J.R. Visual memory task for rats reveals an essential role for hippocampus and perirhinal cortex // PNAS. 2004. V. 101(14). P. 5064–5068. doi: 10.1073/pnas.0308528101.
- Prusky G.T., Harker K.T., Douglas R.M., Whishaw I.Q. Variation in visual acuity within pigmented, and between pigmented and albino rat strains // Behavioural Brain Research. 2002. V. 136. P. 339-348.
- Quirk G.J., Muller R.U., Kubie J.L. The firing of hippocampal place cells in the dark depends on the rat’s recent experience // The Journal of Neuroscience. 1990. V. 10. № 6. P. 2008-2017.
- Riesenhuber M., Poggio T. Neural mechanisms of object recognition // Current Opinion in Neurobiology. 2002. V. 12. P. 162–168.
- Save E., Cressant A., Thinus-Blanc C., Poucet B. Spatial firing of hippocampal place cells in blind rats // The Journal of Neuroscience. 1998. V. 18. № 5. P. 1818-1826.
- Tonkiss J., Shultz P., Galler J.R. Long-Evans and Sprague-Dawley rats differ in their spatial navigation performance during ontogeny and at maturity // Dev. Psychobiol. 1992. V. 25. № 8. P. 567-579.
- Ungerleider L.G., Mishkin M. Two cortical visual systems // Analysis of visual behavior / D.J. Ingle, M.A. Goodale, R.J.W. Mansfield (Eds.). Cambridge, MA: MIT Press. 1982. P. 549–586.
- Vermaercke B., Op de Beeck H. A multivariate approach reveals the behavioral templates underlying visual discrimination in rats // Current Biology. 2012. V. 22. №1. P. 50-55. doi: 10.1016/j.cub.2011.11.041.
- Whishaw I.Q., Hines D.J., Wallace D.G. Dead reckoning (path integration) requires the hippocampal formation: evidence from spontaneous exploration and spatial learning tasks in light (allothetic) and dark (idiothetic) tests // Behavioural Brain Research. 2001. V. 127. P. 49-69.
- Zernicki B. Effects of binocular deprivation and specific experience in cats: behavioural, electrophysiological, and biochemical analysis. // Brain mechanisms in memory and learning: from single neuron to man / Blazier M. (Ed.). New York: Raven Press. 1979. P. 179-195.
Информация об авторах
Метрики
Просмотров web
За все время: 4011
В прошлом месяце: 19
В текущем месяце: 18
Скачиваний PDF
За все время: 757
В прошлом месяце: 4
В текущем месяце: 1
Всего
За все время: 4768
В прошлом месяце: 23
В текущем месяце: 19