Формирование нового элемента опыта на разных этапах онтогенеза: предварительные итоги
Аннотация
Общая информация
Ключевые слова: структура индивидуального опыта, научение, регистрация импульсной активности нейронов, онтогенез
Рубрика издания: Экспериментальные исследования психофизиологической проблемы
Тип материала: материалы конференции
Для цитаты: Соловьева О.А. Формирование нового элемента опыта на разных этапах онтогенеза: предварительные итоги // Экспериментальная психология в России: традиции и перспективы.
Фрагмент статьи
Введение
Одним из вопросов, которые исследуются в психологии, является вопрос о том, как изменяется способность к научению с возрастом. В рамках системно-эволюционного подхода (СЭП), предложенного В. Б. Швырковым на основе теории функциональных систем П. К. Анохина, постулируется, что в процессе научения в виде элемента индивидуального опыта (ЭИО) фиксируется новое соотношение индивида со средой, которое позволяет ему достигнуть полезного приспособительного результата. Слово «фиксация» здесь не случайно, потому что СЭП предполагает, что элементы формируемой на протяжении онтогенеза структуры индивидуального опыта (СИО) обладают онтогенетическим статусом: каждому ЭИО могут быть поставлены в соответствие группы специализированных нейронов, специфически активирующихся при реализации поведенческого акта. Формирование нового ЭИО как «добавки» к существующей СИО рассматривается как специализация новой (а не переспециализация старой) группы нейронов, которые рекрутируются из «запаса» (предположительно, из неспециализированных ранее «молчавших» нейронов) (Gorkin, 1988, Швырков, 1995). На протяжении онтогенеза количество ЭИО растет, таким образом, логично предположить, что «…каждая новая система, сформированная на разных этапах онтогенеза, использует все меньшее число нейронов запаса» (Швырков, 1995, с. 95). На ранних стадиях онтогенеза СИО относительно проста (т.е. содержит мало ЭИО), и «вписывание» нового элемента в нее должно проходить иначе, чем согласование нового ЭИО с более сложной СИО взрослого (и тем более старого индивида) (Александров, 2004). В пользу такого понимания свидетельствуют данные о том, что при одинаковой эффективности обучения разные наборы структур мозга обеспечивают выполнение задания у более молодых и более пожилых людей (Cabeza et al., 2002). У молодых животных помещение в новую обстановку в половине случаев приводит к формированию новых когнитивных карт, в то время как у старых животных с нарушениями памяти они вообще не формируются (Tanila et al., 1997).
Полный текст
Введение
Одним из вопросов, которые исследуются в психологии, является вопрос о том, как изменяется способность к научению с возрастом. В рамках системно-эволюционного подхода (СЭП), предложенного В. Б. Швырковым на основе теории функциональных систем П. К. Анохина, постулируется, что в процессе научения в виде элемента индивидуального опыта (ЭИО) фиксируется новое соотношение индивида со средой, которое позволяет ему достигнуть полезного приспособительного результата. Слово «фиксация» здесь не случайно, потому что СЭП предполагает, что элементы формируемой на протяжении онтогенеза структуры индивидуального опыта (СИО) обладают онтогенетическим статусом: каждому ЭИО могут быть поставлены в соответствие группы специализированных нейронов, специфически активирующихся при реализации поведенческого акта. Формирование нового ЭИО как «добавки» к существующей СИО рассматривается как специализация новой (а не переспециализация старой) группы нейронов, которые рекрутируются из «запаса» (предположительно, из неспециализированных ранее «молчавших» нейронов) (Gorkin, 1988, Швырков, 1995). На протяжении онтогенеза количество ЭИО растет, таким образом, логично предположить, что «…каждая новая система, сформированная на разных этапах онтогенеза, использует все меньшее число нейронов запаса» (Швырков, 1995, с. 95). На ранних стадиях онтогенеза СИО относительно проста (т.е. содержит мало ЭИО), и «вписывание» нового элемента в нее должно проходить иначе, чем согласование нового ЭИО с более сложной СИО взрослого (и тем более старого индивида) (Александров, 2004). В пользу такого понимания свидетельствуют данные о том, что при одинаковой эффективности обучения разные наборы структур мозга обеспечивают выполнение задания у более молодых и более пожилых людей (Cabeza et al., 2002). У молодых животных помещение в новую обстановку в половине случаев приводит к формированию новых когнитивных карт, в то время как у старых животных с нарушениями памяти они вообще не формируются (Tanila et al., 1997).
Теоретическая гипотеза исследования состояла в том, что достижение полезного приспособительного результата (формирование навыка) будет происходить у более молодых индивидов в большей степени за счет формирование новых ЭИО, у старых – за счет изменения отношений между имеющимися ЭИО.
Целью исследования было сравнение СИО на разных стадиях индивидуального развития.
Для проверки гипотезы были сравнены паттерны специализации (соотношение между нейронами, специализированными относительно актов, сформированных до и после попадания в экспериментальную клетку, а также нейронами с неустановленной специализацией) у молодых, взрослых (данные из: Svarnik et al., 2005) и одного старого животного. Часть таких нейронов может быть специализирована относительно актов поведенческого репертуара животных, которые не реализовывались во время проведения данного исследования (например, сексуального поведения), часть – может быть неспециализированными нейронами «запаса», количество которых должно уменьшаться с возрастом по мере приобретения индивидом опыта. Способность к обучению с возрастом снижается (см., напр.: Gallagher, Rapp, 1997), это может быть связано с тем, что у пожилых индивидов уменьшается количество нейронов «запаса», из числа которых рекрутируются нейроны, обеспечивающие выполнение новых поведенческих актов. Поскольку СИО молодых индивидов перед обучением новому навыку состоит из относительно небольшого количества ЭИО и их количество растет с возрастом, можно ожидать, что чем старше будет индивид, тем больше у него будет обнаружено нейронов, специализированных относительно актов, которым он обучился до попадания в экспериментальную обстановку.
Методика
Исследование было проведено на 3 крысах линии Лонг-Эванс (молодых самце и самке 6,5 недель от роду на момент начала обучения и одном старом 24-месячном самце). Животных обучали циклическому инструментальному пищедобывательному поведению в экспериментальной клетке, оборудованной по углам двумя педалями и двумя кормушками. Циклы поведенческих актов на первой в истории обучения стороне экспериментальной клетки формировали поэтапно в течение нескольких дней, на второй стороне – без этапов в течение 1–2 дней за счет «переноса» навыка. Каждый день с животными проводили обучение одному этапу поведения до достижения критерия обученности (5 актов подряд при обучении подходу к кормушке, середине стенки, педали; 10 полных циклов пищедобывательного поведения подряд). Если животное не обучалось акту (или циклу поведения) в течение одной сессии обучения (свыше 50 реализаций акта или цикла), обучение проводили в последующие дни. Во время операции под наркозом обученным животным устанавливали конструкцию для острого погружения стеклянных или вольфрамовых (World Precision Instruments, USA) микроэлектродов с сопротивлением 1,5–3 МОм в заднюю цингулярную область коры (Р: 4,5–5, L: 0,8–1,3) (Paxinos, Watson, 1997).
Через несколько дней после операции начинали регистрировать импульсную активность нейронов во время реализации животными циклического инструментального пищедобывательного поведения на обеих сторонах экспериментальной клетки. На основе актографических отметок выделяли по 4 поведенческих акта на каждой стороне экспериментальной клетки: проверка кормушки и захват пищи, отход от кормушки в сторону педали, нажатие на педаль, отход от педали в сторону кормушки. Активность нейронов оценивали по ее средней частоте в каждом из 8 выделенных актов циклического пищедобывательного поведения крысы в экспериментальной клетке, а также по вероятности наличия активации в акте. За активацию принимали превышение средней частоты в акте не менее чем в полтора раза над «фоном» (частотой разрядов на протяжении сессии регистрации). В случае 100 %-ного наличия активации во всех реализациях определенного поведенческого акта, считали, что нейрон специализирован относительно него. Нейроны классифицировали на нейроны с неустановленной специализацией (НС); нейроны, специализированные относительно актов поведения, сформированных до попадания в экспериментальную клетку («старые», С); нейроны, специализированные относительно актов поведения, которым животное обучилось в экспериментальной клетке («новые», Н). К С-нейронам относили нервные клетки, разряжавшиеся при движении животного в определенном направлении или при поедании пищи в разных условиях – с пола, из рук экспериментатора, из обеих кормушек; к Н-нейронам, – нейроны, активировавшиеся при манипулировании одной или обеими педалями, при захвате пищи из какой-либо одной кормушки.
Для статистического анализа использовали пакет программ SPSS 17.0 (SPSS Inc., USA). Сравнение выборок нейронов с разными специализациями проводили с помощью точного критерия Фишера. Различия считали статистически значимыми при уровне значимости р<0,05.
Результаты и их обсуждение
Для анализа были отобраны 72 нейрона, активность которых была зарегистрирована во время реализации животными не менее чем 10 циклов пищедобывательного поведения на каждой стороне экспериментальной клетки. Три нейрона из 25, зарегистрированных у молодых животных, были отнесены к Н-нейронам, оставшиеся 22 – к НС-нейронам. Из 47 нейронов, зарегистрированных у старой крысы, 5 нейронов были отнесены к Н-нейронам, 11 – к С-нейронам, 31 – к НС-нейронам. Паттерны специализации нейронов у молодых (А), взрослых (Б) (данные из: Svarnik et al., 2005) и старого (В) животного представлены на рисунке 1.
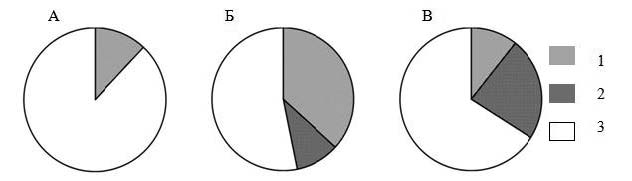
Рис. 1. Паттерны специализации нейронов задней цингулярной коры у животных разного возраста Примечание: А – молодые животные, Б – взрослые (данные из: Svarnik et al., 2005) животные, В – старое животное; 1 – Н-нейроны, 2 – С-нейроны, 3 – НС-нейроны.
Нами было зарегистрировано статистически значимо меньше Н-нейронов (χ² = 5,93, df = 1, p = 0,01) и больше НС-нейронов (χ² = 10,75, df = 1, p = 0,001) у молодых крыс, по сравнению со взрослыми 5–9-месячными животными в опытах В. В. Гаврилова (Svarnik et al., 2005). Мы не зарегистрировали ни одного С-нейрона у молодых животных, в то время как у взрослых животных в задней цингулярной области коры их количество достигает 10% от всех зарегистрированных нейронов (различия между молодыми и взрослыми животными были статистически незначимы, р>0,1). У данного старого животного было зарегистрировано статистически значимо меньше Н-нейронов (χ² = 11,57, df = 1, p = 0,001) и больше С-нейронов (χ² = 5,58, df = 1, p = 0,021), по сравнению со взрослыми животными (Svarnik et al., 2005). У молодых животных было зарегистрировано статистически значимо больше НСнейронов, чем у старого (χ² = 4,08, df = 1, p = 0,037).
Различия в паттернах специализации нейронов, зарегистрированных у старой крысы в данном исследовании и взрослых крыс в опытах В. В. Гаврилова (Svarnik et al., 2005), соответствует предположению, что в старости обучение может происходить в большей степени за счет реорганизации отношений между существующими ЭИО, а не за счет появления новых. Следует отметить, что с возрастом растет вариативность некоторых поведенческих показателей (Morse, 1993), старые индивиды в большей степени отличаются друг от друга, чем взрослые, что дает основание ожидать, что у старых индивидов будет обнаружена значительная вариативность паттерна специализации нейронов, отражающих формирование индивидуальной структуры опыта.
Специализацию части зарегистрированных нейронов установить не удалось, больше всего таких нейронов было найдено у молодых животных. У них встречались нейроны, которые давали специфические активации в актах пищедобывательного поведения, но не со 100 %-ной вероятностью, поэтому они не могли быть отнесены к Н-нейронам. Связь активности этих клеток с поведением у молодых животных могла быть слабее, чем у взрослых, потому что с ними проводили меньше сессий обучения (личное сообщение В.В. Гаврилова), а в первые дни после обучения в задней цингулярной области коры удается зарегистрировать меньше специализированных Н-нейронов, чем спустя неделю или две (Горкин и др., 2004). У старого животного, в отличие от молодых, некоторые НС-нейроны, модулируемые пищедобывательным поведением, активировались при реализации актов комфортного поведения (например, умывания), которым животное обучилось вскоре после рождения. Таким образом, можно предположить, что среди НС-нейронов у молодых индивидов больше клеток, связанных с приобретением любого нового опыта, у старых – обеспечивающих реализацию любых выученных ранее актов (комфортного, сексуального и пр. поведения).
Заключение
При обучении с возрастом количество нейронов, обеспечивающих формирование новых ЭИО, уменьшается по отношению к количеству нейронов, представляющих старые ЭИО.
Литература
-
Александров Ю.И., Греченко Т. Н, Гаврилов В.В. и др. Закономерности формирования и реализации индивидуального опыта // Журнал высшей нервной деятельности. 1997. Т. 47. С. 243–260.
-
Александров Ю. И. Научение и память. Системная перспектива // Вторые симоновские чтения. М.: Изд-во РАН, 2004.
-
Горкин А.Г., Кузина Е.А., Александров Ю.И. Психофизиологические закономерности формирования системной структуры индивидуального опыта в норме и патологии // Первая Российская конференция по когнитивным наукам: Тезисы докладов. Казань.: Изд-во Казан. ун-та, 2004. С. 69–70.
-
Швырков В. Б. Введение в объективную психологию. Нейрональные основы психики. М.: Издво ИП РАН, 1995.
-
Cabeza R., Anderson N. D., Locantore J. K., McIntosh A. R. Aging gracefully: compensatory brain activity in high-performing older adults // NeuroImage. 2002. V. 17. P. 1394–1402.
-
Gallagher M., Rapp P. R. The use of animal models to study the effects of aging on cognition // Ann Rev Psychol. 1997. V. 48. P. 339–370.
-
Gorkin A. G. Learning and neuronal specialization // Psychology of Cognitive Processes / Еds V. B. Shvyrkov et al. M., 1988. Р. 99–104.
-
Morse C. K. Does variability increase with age? An archival study of cognitive measures // Psychol Aging. 1993. V. 8. Р. 156–164.
-
Paxinos G., Watson C. The rat brain in stereotaxic coordinates. The 6th edition. San Diego: Academic, 1997.
-
Svarnik O. E. Alexandrov Yu. I., Gavrilov V. V., Grinchenko Yu. V. Fos expression and task-related neuronal activity in rat cerebral cortex after instrumental learning // Neurosci. 2005. V. 136. P. 33–42.
-
Tanila H., Sipilä P., Shapiro M., Eichenbaum H. Brain aging: impaired coding of novel environmental cues// Journ Neurosci. 1997. V. 17. P. 5167–5174.
Информация об авторах
Метрики
Просмотров web
За все время: 2080
В прошлом месяце: 19
В текущем месяце: 4
Скачиваний PDF
За все время: 54
В прошлом месяце: 2
В текущем месяце: 0
Всего
За все время: 2134
В прошлом месяце: 21
В текущем месяце: 4