Экспериментальная психология
2013. Том 6. № 2. С. 101–114
ISSN: 2072-7593 / 2311-7036 (online)
Модификация плана развертки схемы собственного тела в процессе научения при решении задачи на нахождение обходного пути у улиток вида Achatina fulica *
Аннотация
Общая информация
* Исследование поддержано грантом Президента Российской Федерации № МК-2816.2012.6.
Ключевые слова: психическое отражение, самоотражение, отражение внешней среды, филогенез психики, моллюски, улитка
Рубрика издания: Эволюционная и сравнительная психология
Тип материала: научная статья
Для цитаты: Хватов И.А., Харитонов А.Н. Модификация плана развертки схемы собственного тела в процессе научения при решении задачи на нахождение обходного пути у улиток вида Achatina fulica // Экспериментальная психология. 2013. Том 6. № 2. С. 101–114.
Полный текст
Изучение специфики самоотражения животных разных таксономических групп является перспективной задачей зоопсихологии и сравнительной психологии. За последние несколько лет авторами настоящей статьи был проведен ряд эмпирических исследований, позволивших выявить особенности самоотражения различных позвоночных и беспозвоночных (Филиппова, Хватов, 2011; Хватов, 2010 а; 2010 б; 2011 а; 2011 б; Хватов, Харитонов, 2012). В частности, в результате исследования представителей семейства легочных брюхоногих моллюсков – улиток-ахатинид вида Achatina fulica – было установлено, что эти животные способны отражать физические границы и размеры наружной мягкой части собственного тела (ноги и головы) и учитывать их при ориентации во внешнем пространстве, но не способны учитывать физические границы и размеры собственной раковины (Хватов, Харитонов, 2012). Результаты анализа полученных в ходе эксперимента данных свидетельствовали об отсутствии сколь-нибудь заметных признаков выработки улитками навыка учета физических границ собственной раковины, а следовательно, об отсутствии у них способности к модификации содержания самоотражения. Подобный результат связан, с нашей точки зрения, с условиями эксперимента: временной интервал между экспериментальными пробами, проведенными с каждым отдельным животным, был довольно большим (две пробы в неделю).
Под самоотражением мы понимаем процесс и результат отражения субъектом своей внутренней объективной реальности: характеристик своего организма – в контексте данного исследования речь будет идти о размерах и границах собственного тела (Хватов, 2010 а).
Между тем, существует немало свидетельств способности брюхоногих моллюсков к научению. Самой простой формой последнего следует считать привыкание: ослабление ответной реакции на ритмическую тактильную стимуляцию у улиток семейства гелицид – Helix pomatia (Балабан, Захаров, 1992) и H. albilabris при ритмическом покачивании (Шовен, 2009), а также привыкание и предпочтение определенного вида пищи у A. fulica (Chase, Croll, Zeichner, 1980), виноградной улитки H. pomatia (Teyke, 1995) и прудовика обыкновенного Lymnaea stagnalis (Бондаренко, Боричева, 2002).
Более сложной формой является ассоциативное научение, способность к которому была исследована: у виноградной улитки H. pomatia посредством проверки способности к выработке оборонительной реакции в ответ на пищевой раздражитель (Балабан, Захаров, 1992); у улиток-физ вида Physa gyrina – посредством проверки способности совершать жевательные движения в ответ на прикосновения (Шовен, 2009); у слизней Limax maximus – посредством проверки способности к модификации пищевых предпочтений при негативной стимуляции горького вкуса сульфата хинидина (Sahley, Gelperin, Rudy, 1981).
Улитки A. fulica могут запоминать расположение источников пищи и возвращаться к ним. Молодые особи более подвижны и преодолевают большие расстояния в течение дня, а также способны к дальним миграциям. Обычно для отдыха в одно и то же место они не возвращаются. У старых же улиток, напротив, имеется место, где они предпочитают отдыхать и откуда выходят на поиски питания, не удаляясь от него более чем на 5 метров. При переносе улитки в место отдыха другой особи A. fulica (в пределах 30 метров) она всё равно возвращается к своему прежнему месту обитания (Tomiyama, 1992). При возвращении в место своего обитания (хоуминге), а также при ориентации во внешнем пространстве в целом улитки способны определять направление движения по слизи, оставленной при перемещении другими представителями своей популяции, что позволяет им быстрее находить правильный маршрут (Rollo, Wellington, 1981). Более детальное изучение феномена хоуминга было проведено также в исследованиях поведенческих реакций морских брюхоногих моллюсков приливно-отливной зоны – блюдечек Patella granularis (Шовен, 2009).
Отдельного внимания заслуживают исследования способности к научению моллюсков ориентироваться в лабиринте. В частности, Гарт и Митчелл (Garth, Mitchell, 1926) сформировали у хищной сухопутной улитки Rumina decollata навык прохождения Т-образного лабиринта, основными признаками выработки которого являлись: сокращение времени решения экспериментальной задачи, уменьшение количества ошибок, а также выпрямление траектории движения моллюска в лабиринте. В качестве стимуляции использовался дневной и электрический свет (данный вид улиток характеризуется отрицательным фототаксисом), в качестве отрицательного подкрепления – воздействие нагретой проволокой.
Исследования координации движений моллюсков (в частности, ее психофизиологической регуляции) были проведены главным образом на материале изучения поведения головоногих – наиболее высокоорганизованных представителей этой группы, и в первую очередь, осьминогов. Данный вид головоногих обладает сложно устроенными эффекторными органами, позволяющими осуществлять тонкие локомоторные и манипуляционные операции (Рупперт и др., 2008). Регуляция моторики этих головоногих имеет сложную иерархическую организацию, мозг интегрирует огромное количество тактильных и визуальных сигналов (Zullo et al., 2009). Осьминоги, обладая, казалось бы, неограниченным числом степеней свободы в движениях щупальцами, для выполнения отдельных операций способны создавать квазисочлененные конструкции, подобные конечностям позвоночных (Sumbre et al., 2006). Однако результаты других исследований показали, что осьминоги способны проникать отдельным щупальцем в лабиринт, состоящий из трех отсеков и крепящийся сверху на их аквариум, и доставать приманку, находящуюся в одном из отсеков. Авторы данного исследования предположили, что животные осуществляли регуляцию собственных движений с помощью обработки зрительной информации (Gutnick et al., 2011).
Подводя итог вышеизложенному, можно сделать вывод о целесообразности дальнейшего более детального исследования способности брюхоногих моллюсков модифицировать самоотражение (схему собственного тела) посредством научения. Результаты проведенных нами ранее исследований указывают на наличие у улиток вида A. fulica способности к изменению конфигурации мягкого тела при решении задачи проникновения в отверстия разного размера (Хватов, Харитонов, 2012). Наличие такой способности служит основанием для экспериментального исследования самоотражения брюхоногих моллюсков в условиях необходимости гибкой регуляции конфигурации тела при решении более сложных и разнотипных задач.
Таким образом, цель настоящего исследования состоит в выявлении способности улитки вида Achatina fulica модифицировать план развертки схемы собственного тела в процессе научения при решении задачи на нахождение обходного пути.
Методика экспериментального исследования
Характеристики экспериментальных животных: 40 улиток вида A. fulica, высота раковины 30–35 м; особи содержались отдельно друг от друга (для исключения возможности их взаимовлияния как побочной переменной).
Оборудование. В соответствии с основной задачей исследования, заключающейся в изучении способности улитки двигать именно мягкой частью собственного тела для достижения приманки, были созданы такие экспериментальные условия, которые ограничивали свободу локомоции улитки, направленную на перемещение всего тела. Пусковая камера 75×45 мм была сконструирована таким образом, чтобы выходное отверстие в виде щели высотой 15 мм и шириной 45 мм могло бы пропустить лишь мягкую часть тела моллюска (голова и нога), но не раковину. Сразу за выходным отверстием на расстоянии 20 мм от камеры располагалась преграда в виде металлической сетки с отверстиями диаметром 0,5 мм, за которой помещалась приманка (кусочек огурца). Запах приманки свободно проникал через отверстия в сетке (при пищевом поведении улитки ориентируются на вкус и запах объекта) – то есть само устройство экспериментальной установки предполагало, что для решения задачи (достижения приманки) улитки должны были бы обогнуть преграду таким образом, чтобы при перемещении тела в сторону приманки их раковина не покидала пусковой камеры.
Фиксируемые показатели. В качестве независимой переменной выступала преграда из сетки, отделявшая животных от приманки. В качестве зависимых переменных выступали: а) время решения экспериментальной задачи (от момента помещения испытуемого внутрь пусковой камеры до достижения приманки), б) виды двигательных операций, совершаемых улитками внутри экспериментальной установки, на основании сочетаний которых были далее составлены этограммы.
План эксперимента. В соответствии с целями и задачами эксперимента из всей выборки улиток нами было выделено две группы: экспериментальная – 30 улиток, в экспериментальной установке которых наличествовала преграда из сетки; контрольная выборка – 10 улиток, в экспериментальной установке которых отсутствовала преграда из сетки. Сравнительный анализ результатов выполнения экспериментального задания в двух группах животных позволяет установить влияние независимой переменной на поведение испытуемых, а также ответить на вопрос, насколько быстро моллюски способны решить экспериментальную задачу при отсутствии преграды.

Рис. 1, а. Фотография экспериментальной установки (крышка, закрывающая пусковую камеру снята): а – пусковая камера, б – щелевое отверстие в стенке пусковой камеры, в – преграда из сетки
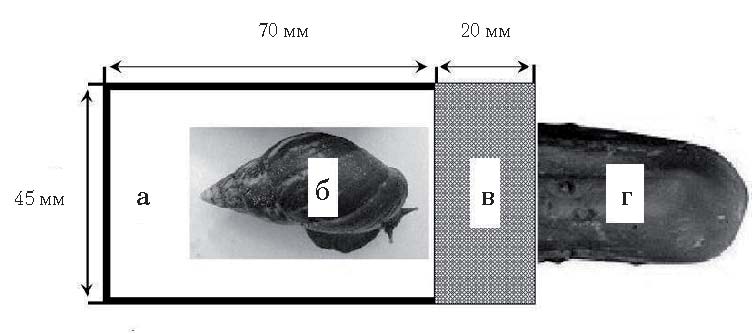
Рис 1, б. Схема экспериментальной установки (вид сверху): а – пусковая камера, б – улитка, в – преграда из сетки, г – приманка
Исследование процесса научения предполагает в первую очередь выявление динамики изменения фиксируемых показателей: времени решения экспериментальной задачи и поведенческой активности животных из обеих выборок. В качестве эмпирического критерия, подтверждающего факт научения, выступили достоверные показатели сокращения времени решения задачи и упрощение схемы поведения (этограммы) у животных экспериментальной выборки к концу эксперимента по сравнению с его началом.
Каждая улитка участвовала в 30 экспериментальных пробах. Пробы шли сразу одна за другой в течение одного дня. Перед началом эксперимента для обеспечения необходимой мотивации животные подвергались пищевой депривации: животных не кормили 10 суток. Кроме того, в случае положительного результата – формирования у животных навыка нахождения обходного пути – предполагалось проведение дополнительной состоящей из 30 проб экспериментальной серии с теми же животными спустя десять суток для уточнения вопроса о том, сохраняется ли у них ранее сформированный навык.
Результаты анализа временных показателей решения экспериментальной задачи
Общая динамика временных интервалов решения экспериментальной задачи животными экспериментальной и контрольной групп представлена на рис. 2.
Как видно из графика, у испытуемых экспериментальной выборки время решения экспериментальной задачи резко сокращается от 1-й пробы (в среднем 27,5 мин., SD=8,4) к 12-й (в среднем 8 мин., SD=4,2), затем темп сокращения временных интервалов снижается и к 30 пробе время решения экспериментальной задачи в среднем составляет 4,3 мин. (SD=2,8).
С помощью ANOVA Фридмана была выполнена проверка нулевой статистической гипотезы (H0) об отсутствии достоверных отличий между эмпирическими распределениями временных интервалов, затраченных испытуемыми экспериментальной выборки на решение экспериментальной задачи, на каждой из 30 проб. Полученные результаты – ANOVA χ2=395,8589 p<0,01 (N=30, df=29) – привели к отвержению нулевой гипотезы H0 и принятию гипотезы H1: эмпирические распределения значений времени, затраченного животными экспериментальной группы на выполнение каждой из 30 проб, достоверно отличаются друг от друга.
Что касается контрольной выборки, то, как видно из графика на рис. 2, на протяжении всего эксперимента время решения экспериментальной задачи практически не изменялось: 1-я проба – среднее время 2,5 мин. (SD=0,6), 15-я проба – среднее время 2,4 мин. (SD=0,5), 30-я проба – среднее время 2,4 мин. (SD=0,6).
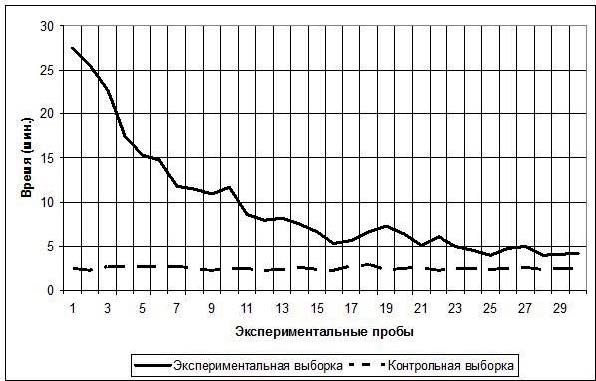
Рис. 2. Среднее время, затраченное испытуемыми обеих выборок, на решение экспериментальной задачи в 30 экспериментальных пробах
С помощью ANOVA Фридмана была выполнена проверка нулевой статистической гипотезы (H0) об отсутствии достоверных отличий между эмпирическими распределениями значений времени, затраченного животными контрольной выборки на решение экспериментальной задачи, на каждой из 30 проб. Результат ANOVA χ2=12,52534 p=0,99 (N=30, df=29) свидетельствует, что гипотеза H0 подтвердилась.
Далее с помощью критерия χ2 была выполнена проверка нулевой статистической гипотезы (H0) об отсутствии достоверных отличий между эмпирическими распределениями средних значений времени, затрачиваемого на решение экспериментальной задачи животными экспериментальной и контрольной выборок. Полученный результат χ2=1060,046 p<0,01 (df=29) привел к отвержению гипотезы H0 и принятию H1: эмпирические распределения средних значений времени, затрачиваемого на решение экспериментальной задачи животными экспериментальной и контрольной выборок, достоверно отличаются друг от друга.
Была также проведена дополнительная экспериментальная серия из 30 проб с животными экспериментальной группы спустя 10 дней после первой серии; дополнительная серия была проведена с целью проверки способности улиток сохранить ранее сформированный навык. Результаты приведены на рис. 3.
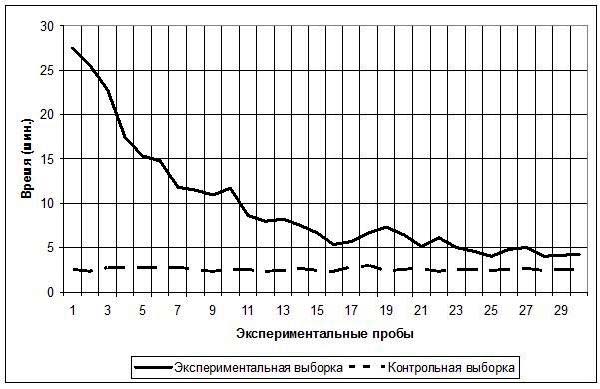
Рис. 3. Среднее время, затраченное испытуемыми экспериментальной выборки, на решение экспериментальной задачи в 30 экспериментальных пробах в первой и второй экспериментальных сериях
Как видно из графиков, начало второй серии характеризуется снижением средних показателей времени, затраченного животными экспериментальной группы на выполнение задачи, по сравнению с теми же показателями первой серии. Усредненные результаты по 30 испытуемым в первой пробе: серия 1 – 27,5 мин. (SD=8,4), серия 2 – 16,5 мин. (SD=3,2). Во второй серии также наблюдается динамика быстрого снижения времени решения экспериментальной задачи, а после 15-й пробы в обеих сериях эксперимента происходит уравнивание показателей времени решения задачи, о чем свидетельствуют усредненные результаты по 30 испытуемым: так, в 15-й пробе: серия 1 – 6,6 мин. (SD=3,1), серия 2 – 5,8 мин. (SD=1,6); в 30-й пробе: серия 1 – 4,3 мин. (SD=2,8), серия 2 – 4,5 мин. (SD=2).
С помощью критерия χ2 была выполнена проверка нулевой статистической гипотезы H0 об отсутствии достоверных отличий между эмпирическими распределениями среднего значения времени, затраченного на решение задачи поиска обходного пути животными экспериментальной выборки в первой и второй экспериментальных сериях. Результат: χ2=46,00448 p=0,02 (df=29). Таким образом, H0 была отвергнута. Однако и гипотеза о наличии достоверных отличий H1 также не могла быть принята, поскольку значения критерия попадали в зону неопределенности.
В связи с тем, что основные отличия показателей между двумя экспериментальными сериями приходились на первые 15 проб, мы отдельно сопоставили между собой эмпирические распределения средних значений времени, затраченного на решение задачи поиска обходного пути животными экспериментальной выборки, в первой половине (пробы с 1-й по 15-ю) обеих экспериментальных серий и во второй их половине (пробы с 16-й по 30-ю). Результаты сравнения временных показателей в пробах с 1-й по 15-ю обеих экспериментальных серий: χ2=44,05916 p<0,01 (df=14) свидетельствуют в пользу принятия гипотезы H1: эмпирические распределения средних значений времени решения задачи в первых 15 пробах обеих экспериментальных серий достоверно отличаются друг от друга. Результаты по второй половине двух экспериментальных серий: χ2=1,945313 p=0,99 (df=14) – свидетельствуют в пользу принятия гипотезы H0: достоверные отличия между двумя экспериментальными сериями в средних значениях времени, затраченного на решение задачи поиска обходного пути, в последних 15 пробах отсутствуют.
Поведенческая активность улиток
Все многообразие поведенческой активности животных экспериментальной выборки было обобщено в несколько основных видов движений (двигательных операций); затем на основании специфики сочетаний выделенных видов движений были составлены этограммы животных. Для демонстрации качественных изменений в поведении улиток при решении задачи поиска обходного пути были сопоставлены между собой показатели поведенческой активности животных в первых пяти и последних пяти пробах первой экспериментальной серии. Предварительно отметим, что классификации повергались только формы поведенческой активности животных за пределами пусковой камеры; поведение внутри камеры было описано в общих чертах и затем включено в итоговый анализ.
Первым движением, совершаемым всеми животными, являлся выход из пусковой камеры – проникновение головой и ногой в щелевое отверстие. После этого чаще всего (в 86 % случаев за первые пять проб с животными экспериментальной группы, рис. 4) моллюски двигались напрямую к приманке и упирались в сетку (контакт с сеткой): они прижимались к сетке, совершали «жевательные движения» радулой, проникали щупальцами в отверстия в сетке. Лишь в 5 % случаев улитки направлялись непосредственно к приманке, огибая сетку мягким телом (достижение приманки). Кроме того, в 9 % случаев сразу после выхода из пусковой камеры животные вытягивали голову вверх (сюда же относились случаи, когда они проникали в щелевое отверстие в перевернутом состоянии, заползая туда с потолка пусковой камеры через переднюю стенку).
После контакта с сеткой и тщетных попыток добраться до приманки сквозь нее в большинстве случаев (67 % случаев) улитки втягивали тело обратно (втягивание тела – рис. 5), т. е. возвращались обратно в пусковую камеру, где затем совершали различные движения – чаще всего улитки двигались по кругу (по полу, по стенам или же по полу и по потолку) внутри камеры и затем снова проникали в щелевое отверстие. Аналогичное поведение наблюдалось и в случаях, когда улитки после выхода из пусковой камеры сразу вытягивали голову вверх – в 83 % случаев они затем втягивали тело, а также и в случаях, когда после контакта с сеткой моллюски вытягивали голову вверх, прижимая тело к сетке (10 % случаев): после этого они в 86 % случаев втягивали тело обратно.
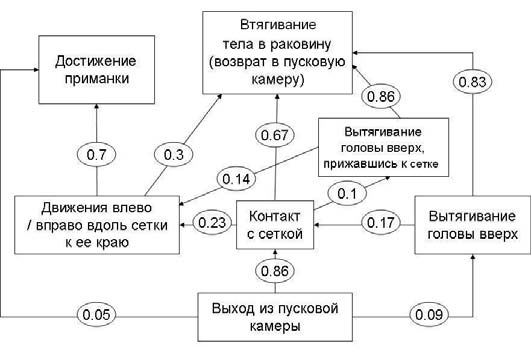
Рис. 4. Этограмма улиток экспериментальной выборки на первых пяти пробах первой экспериментальной серии

Рис. 5. Операция «Втягивание тела в раковину» после контакта с сеткой
В 23 % случаев после контакта с сеткой животные двигались вправо или влево вдоль сетки (соприкасаясь с ней) к ее краю, т. е. двигались по обходному пути к приманке (рис. 6), однако даже и после этого в 30 % случаев моллюски втягивали тело назад, однако в 70 % случаев они достигали приманки.

Рис. 6. Операции «Движение вдоль сетки к ее краю» и «Достижение приманки»
Сравним далее вышеописанное поведение улиток экспериментальной группы в первых пяти пробах первой экспериментальной серии с поведением этих же улиток в последних пяти пробах (с 26-й по 30-ю) первой экспериментальной серии. Этограмма улиток на последних пяти пробах изображена на рис. 7.
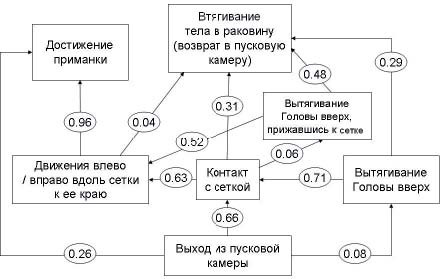
Рис. 7. Этограмма улиток экспериментальной выборки на последних пяти пробах первой экспериментальной серии
Как видно из схемы, к концу экспериментальной серии поведение животных существенно меняется. Во-первых, увеличивается количество прямых движений по обходному пути к приманке даже без соприкосновения с сеткой: в 26 % случаев против 5 % в первых 5 пробах. Во-вторых, количество обходных движений, следующих за контактом с сеткой, также возрастает: 63 % случаев против 23 % в первых 5 пробах. В-третьих, в целом уменьшается доля обратных втягиваний тела, следующих за различными другими операциями: после вытягивания головы вверх (29 % случаев в конце экспериментальной серии против 83 % в начале), после контакта с сеткой (31 % случаев в конце против 67 % в начале), после вытягивания головы вверх в положении «прижавшись к сетке» (48 % случаев в конце против 86 % в начале), после движений вдоль сетки к ее краю (4 % случаев в конце против 30 % в начале).
Далее с применением критерия χ2 проверялась нулевая статистическая гипотеза H0 об отсутствии достоверных отличий между распределениями количества двигательных операций, следующих за выходом из пусковой камеры (достижение приманки, контакт с сеткой, вытягивание головы вверх) у животных экспериментальной группы в первых пяти пробах (с 1-й по 5-ю) и последних пяти (с 26-й по 30-ю) пробах первой экспериментальной серии. Результаты (рис. 8) χ2=7588,293 p<0,01 (df=2) свидетельствовали против гипотезы H0 и принятия как подтвержденной обратной ей гипотезы H1: распределения количества двигательных операций, следующих за выходом из пусковой камеры, у животных экспериментальной группы в первых пяти пробах и последних пяти пробах первой экспериментальной серии достоверно отличаются друг от друга.
Затем с использованием критерия χ2 была проверена нулевая статистическая гипотеза H0 об отсутствии достоверных отличий между распределениями количества двигательных операций, следующих за контактом с сеткой (вытягивание головы вверх, прижавшись к сетке, сворачивание тела в раковину, движение вдоль сетки к ее краю) у животных экспериментальной группы в первых пяти пробах (с 1-й по 5-ю) и последних пяти (с 26-й по 30-ю) пробах первой экспериментальной серии. Результаты (рис. 9) – χ2=13968,44 p<0,01 (df=2) – указывают на то, что гипотеза H0 не нашла подтверждения, в связи с чем принимается гипотеза H1: распределения количества двигательных операций, следующих за выходом из пусковой камеры, у животных экспериментальной группы в первых пяти пробах и последних пяти пробах первой экспериментальной серии достоверно отличаются друг от друга.
Следует отдельно отметить, что на более поздних этапах эксперимента улитки решали экспериментальную задачу с помощью меньшего числа двигательных операций, т. е. их поведение упрощалось.
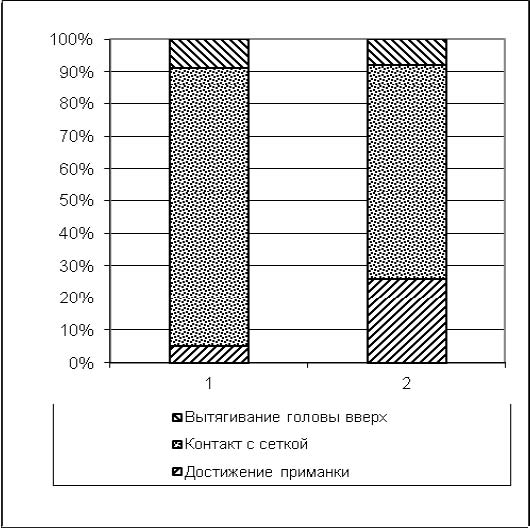
Рис. 8. Суммарное количество двигательных операций всех испытуемых экспериментальной выборки (обозначены числами в гистограмме) в 1-й экспериментальной серии, совершенных после выхода из пусковой камеры: 1 – пробы с 1-й по 5-ю; 2 – пробы с 26-й по 30-ю
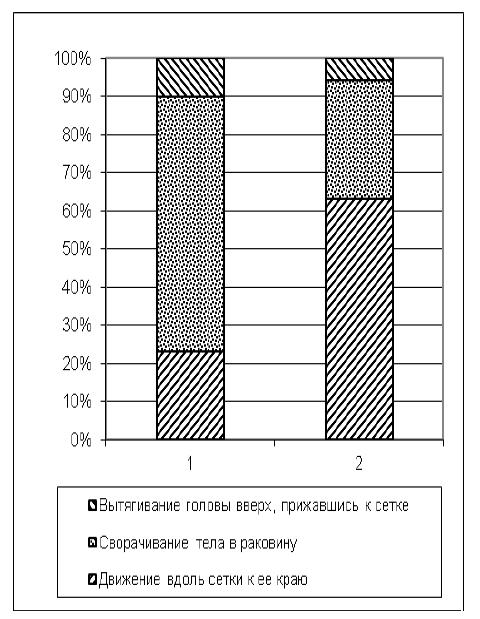
Рис. 9. Суммарное количество двигательных операций всех испытуемых экспериментальной выборки (обозначены числами в гистограмме) в 1-й экспериментальной серии, совершенных после контакта с сеткой: 1 – пробы с 1-й по 5-ю; 2 – пробы с 26-й по 30-ю
Что касается второй экспериментальной серии, проводившейся спустя 10 дней после первой, то динамика изменений поведения животных экспериментальной группы была аналогичной динамике поведенческих изменений первой серии за тем исключением, что общее число двигательных операций, совершенных улитками при решении задачи поиска обходного пути, было пропорционально меньше числа операций, совершенных улитками в ходе первой экспериментальной серии.
Поведение животных контрольной группы было весьма простым: сразу после выхода из пусковой камеры они напрямую направлялись к приманке и достигали ее. Обратные втягивания тела отсутствовали.
Анализ и интерпретация данных
Анализ динамики сокращения времени решения задачи поиска обходного пути в ходе первой экспериментальной серии дает основание утверждать, что у моллюсков формируется навык нахождения обходного пути и достижения приманки. Очевидно, навык складывается за первые 12–15 проб, поскольку затем время решения задачи сокращается весьма медленно. Сравнение этих данных с результатами контрольной группы свидетельствует, что преграда из сетки действительно влияет на поведение улиток – является фактором, затрудняющим достижение приманки.
Сравнение результатов первой и второй экспериментальных серий не позволяет сделать однозначного вывода. Однако поскольку, как отмечалось ранее, основной период формирования навыка – первые 15 проб, мы сосредоточили внимание именно на первой половине двух экспериментальных серий. Здесь отличия существенны: с самого начала второй экспериментальной серии животные не только тратили значительно меньше времени на решение задачи, но также быстро снижалось количество производимых ими двигательных операций. Таким образом, можно утверждать, что ранее сформированное научение сохраняется в памяти улиток в течение 10 дней, поскольку повторно навык формируется значительно быстрее. По-видимому, можно говорить о восстановлении навыка.
Данные о поведении животных экспериментальной группы в начале первой экспериментальной серии свидетельствуют, что у улиток весьма часто встречается движение обратного втягивания тела после контакта с преградой. Очевидно, в данном случае мы имеем дело с инстинктивно детерминированным планом развертки схемы собственного тела: в ответ на контакт с непреодолимой преградой – сеткой – улитка втягивает тело в раковину (частично или полностью) и затем начинает как бы «с нуля» двигаться в совершенно ином направлении. В ходе эксперимента после втягивания тела животные поворачивали назад и начинали двигаться внутри пусковой камеры. Однако такой способ поведения был неэффективен – улитки в любом случае находили обходной путь, хотя первоначально совершали большое количество нерезультативных движений.
К концу первой экспериментальной серии (на последних пяти пробах) поведение животных существенно менялось: помимо количественных изменений (уменьшение количества движений, затрачиваемых на решение поставленной задачи) происходили и значительные качественные изменения двигательных операций. Теперь в ответ на контакт с преградой улитки значительно реже втягивали тело в раковину, а вместо этого двигались вдоль сетки (вправо или влево) к ее краю, т. е. совершали эффективные действия, ведущие к достижению приманки. Даже в случае втягивания тела в раковину улитки чаще всего не поворачивали назад, совершая движения внутри пусковой камеры, а тут же снова проникали через щелевое отверстие, перемещая свое тело в ином направлении, нежели то, где они до этого столкнулись с преградой.
Таким образом, мы можем констатировать, что в ходе нашего эксперимента у животных экспериментальной группы посредствам научения модифицировался план развертки схемы собственного тела в ответ на контакт с преградой. В начале эксперимента он имел следующий вид: контакт с преградой → втягивание тела в раковину → движение всем телом в сторону от преграды. К концу экспериментальной серии план развертки схемы собственного тела приобрел иной вид: контакт с преградой → движение наружной (мягкой) частью собственного тела вдоль преграды → достижение края преграды, преодоление преграды и достижение приманки. Согласно теории самоотражения (Хватов, 2010a), план развертки схемы собственного тела является одним из трех (наравне со схемой собственного тела и перцептивным строем) компонентов психической структуры самоотражения. Соответственно, в ходе эксперимента посредством научения была модифицирована специфика самоотражения улиток вида Achatina fulica. Тот факт, что представители этого вида обладают психической структурой самоотражения наружной (мягкой) части собственного тела и способны учитывать физические границы собственного тела при ориентации во внешнем пространстве, был продемонстрирован результатами проведенных ранее исследований (Хватов, Харитонов, 2012).
Выводы
На основании проведенного анализа результатов данного исследования мы можем констатировать следующее.
-
В организованных экспериментальных условиях для решения задачи поиска обходного пути улитки посредством научения способны не только модифицировать план развертки собственного тела, что выражается в сокращении времени решения данной задачи, но также произвести качественные изменения в способе своего поведения внутри экспериментальной установки;
-
Следы сформированного научения сохраняются в памяти моллюсков как минимум 10 суток, что подтверждается более быстрым формированием этого же навыка (его восстановлением) в аналогичных экспериментальных условиях.
Таким образом, перспективным направлением исследований является проведение аналогичных сравнительных экспериментов по изучению поведенческой активности и поэтапного формирования двигательных навыков у животных, относящихся как к более высоким, так и к более низким стадиям развития психики.
Литература
- Балабан П. М., Захаров И. С. Обучение и развитие: общая основа двух явлений. М.: Наука, 1992.
- Бондаренко В. Ф., Боричева Е. С. Формирование и сохранение памяти на хемосенсорно выделяемые признаки растительной пищи у прудовика обыкновенного Lymnaea stagnalis // Сенсорные системы. 2002. Т. 16. № 2. С.160–163.
- Рупперт Э. Э., Фокс Р. С., Барнс Р. Д. Зоология беспозвоночных: Функциональные и эволюционные аспекты: учебник для вузов: В 4 т. (Перевод с англ. Т. А. Ганф, Н. В. Ленцман, Е. В. Сабанеевой / Под ред. А. А. Добровольской, А. И. Грановича. М.: Академия, 2008. Т. 2.
- Филиппова Г. Г., Хватов И. А. Специфика экспериментального метода в зоопсихологии на примере исследования самоотражения у животных на интеллектуальной стадии развития психики // Современная экспериментальная психология: В 2 т. / Под ред. В. А. Барабанщикова. М.: Изд. «Институт психологии РАН», 2011. Т. 1. С. 499–511.
- Хватов И. А. Особенности самоотражения у животных на разных стадиях филогенеза: Дисс. … канд. психол. наук. М., 2010 а.
- Хватов И. А. Эмпирическое исследование проблемы филогенетических предпосылок становления самосознания // Знание. Понимание. Умение. 2010 б. № 2. С. 242–247.
- Хватов И. А. Специфика самоотражения у вида Periplaneta americana // Экспериментальная психология. 2011 а. № 1. C. 28–40.
- Хватов И. А. Эмпирическое исследование восприятия самих себя у понгид (на примере рисования перед зеркалом у орангутанов) // 125 лет Московскому психологическому обществу: Юбилейный сборник РПО: В 4-х т. / Отв. ред. Д. Б. Богоявленская, Ю. П. Зинченко. М.: МАКС Пресс, 2011 б. Т. 2. С. 46–47.
- Хватов И. А., Харитонов А. Н. Специфика самоотражения у вида Achatina fulica // Экспериментальная психология. 2012. № 3. C. 96–107.
- Шовен Р. Поведение животных: пер. с фр. / Под ред. и с предисл. Л. В. Крушинского. Изд. 2-е. М.: Либроком, 2009.
- Chase R., Croll R. P., Zeichner Z. Z. Aggregation in snails, Achatina fulica // Behavioral and Neural Biology. 1980. V. 30. № 2. P. 218–230.
- Garth T. R., Mitchell M. P. The learning curve of a land snail // J. of Comp. Psych. 1926. V. 6. № 1. P. 103–113. doi: 10.1037/h0072322
- Gutnick T., Byrne R. A., Hochne B., Kuba M. Octopus vulgaris uses visual information to determine the location of its arm // Current Biology. 2011. V. 21. P. 1–3. doi:10.1016/j.cub.2011 .01.0 52
- Rollo С. D., Wellington W. G. Environmental orientation by terrestrial Mollusca with particular reference to homing behaviour // Canadian Journal of Zoology. 1981. V. 59. № 2. P. 225–239.
- Sahley C., Gelperin A., Rudy J. W. One-trial associative learning modifies food odor preferences of a terrestrial mollusk // PNAS. 1981. V. 78. № 1. P. 640–642.
- Sumbre G., Fiorito G., Flash T., Hochner B. Octopuses use a human-like strategy to control precise pointto-point arm movements // Current Biology. 2006. V. 16. №. 8. P. 767–772. doi:10.1016/j.cub.2006.02.069
- Teyke T. Food attraction conditioning in the snail Helix pomatia // Comp. Physiol. A. 1995. V. 177. № 4. P. 409–414.
- Tomiyama К. Homing behaviour of the giant African snail, Achatina fulica // Journal of Ethology. 1992. V. 10. № 2. P. 139–146.
- Zullo L., Sumbre G., Agnisola C., Flash T., Hochner B. Nonsomatotopic Organization of the Higher Motor Centers in Octopus // Current Biology. 2009. V. 19. № 19. P. 1632–1636. doi:10.1016/j.cub.2009.07.067.
Информация об авторах
Метрики
Просмотров web
За все время: 3128
В прошлом месяце: 20
В текущем месяце: 7
Скачиваний PDF
За все время: 744
В прошлом месяце: 5
В текущем месяце: 1
Всего
За все время: 3872
В прошлом месяце: 25
В текущем месяце: 8