Клиническая и специальная психология
2022. Том 11. № 1. С. 192–209
doi:10.17759/cpse.2022110109
ISSN: 2304-0394 (online)
Зрительный прайминг у детей с расстройствами аутистического спектра
Аннотация
Общая информация
Ключевые слова: зрительный прайминг, дошкольный возраст, расстройства аутистического спектра, восприятие
Рубрика издания: Эмпирические исследования
Тип материала: научная статья
DOI: https://doi.org/10.17759/cpse.2022110109
Финансирование. Исследование выполнено при финансовой поддержке Российского фонда фундаментальных исследований (РФФИ) в рамках научного проекта № 17-06-00644-ОГН
Благодарности. Авторы благодарят за помощь в организации работы коллектив с Санкт-Петербургского «Центра реабилитации ребенка»
Получена: 29.06.2021
Принята в печать:
Для цитаты: Черенкова Л.В., Соколова Л.В. Зрительный прайминг у детей с расстройствами аутистического спектра [Электронный ресурс] // Клиническая и специальная психология. 2022. Том 11. № 1. С. 192–209. DOI: 10.17759/cpse.2022110109
Полный текст
Введение
В настоящее время не вызывает сомнения наличие особого типа
восприятия
у людей с расстройством аутистического спектра (РАС) [Haigh, 2018]. Людям с РАС
характерны фрагментарность восприятия и повышенное внимание к отдельным
несущественным для общего контекста элементам зрительной сцены, что затрудняет
интеграцию этих элементов в единый репрезентативный образ [Robertson, 2017]. Формирование
репрезентативного образа обычно базируется на иерархически организованном
взаимодействии процессов обработки зрительной информации на разных уровнях
нервной системы [Kimchi, 2015]. При аутизме и РАС наблюдается иной тип интеграции,
который проявляется уже на нижних уровнях нервной системы, где выделяются
и взаимодействуют элементарные признаки воспринимаемых объектов. Так,
в одной из работ было высказано предположение о том, что при аутизме более
развиты системы, ответственные за переработку высокочастотной пространственной
информации по сравнению с низкочастотной [Sutherland, 2010]. Однако результаты другого
исследования показывают, что восприятие глобальных образов, содержащих низкие
пространственные частоты, возможно и при аутизме, если к ним привлекать
внимание испытуемых [Van der Hallen, 2016]. Это направило исследователей на изучение процессов
сенсорного внимания, так как именно механизмы распределения и переключения
внимания обусловливают накопление сенсорного опыта, необходимого для процесса
репрезентации [Kristjansson, 2019; Shomstein, 2012]. При РАС отмечают особый характер организации функции
внимания и механизмов его мозгового обеспечения [Amso, 2014]. Однако характеристики
«антиципирующего внимания», связанного с функцией распределения внимания между
отдельными элементами текущей и упреждающей информации, поступающей
последовательно во времени, при аутизме и РАС изучены мало.
Одновременно исследовать характеристики процессов интеграции
информации
и сенсорного внимания позволяет процедура прайминга. Зрительный прайминг
представляет собой изменение способности идентифицировать, воспроизводить или
классифицировать текущие объекты под влиянием восприятия уже предъявлявшейся
прежде зрительной информации [Janiszewski, 2014]. Под влиянием этой информации могут
изменяться показатели успешности и скорости реакции на текущие стимулы в
зависимости от их сходства с предваряющей информацией (праймом). При полном
подобии прайма и целевых стимулов (повторный прайминг) параметры реакции
улучшаются, а при несоответствии прайма целевым объектам скорость и точность
реакции ухудшаются (отрицательный прайминг) [Janiszewski, 2014]. Когда прайм и целевой стимул
отличаются по отдельным параметрам, знак и величина изменения реакции
(прайминг-эффект) будут зависеть от наличия общих признаков в их составе, таких
как цвет, пространственное положение, ориентация, отдельные локальные элементы,
общая конфигурация [Janiszewski, 2014; Kristjansson, 2019; McLaughlin, 2021; Mo, 2019; Pokhoday, 2019]. Выявление характеристик изменения
параметров реакции под влиянием прайма способствует определению тех признаков
объектов, которые являются значимыми при обработке и отборе информации,
необходимой для реализации деятельности. Особый случай представляет изучение
эффекта прайминга при изменении времени задержки между праймом и целевым
стимулом в пределах сотен миллисекунд, в течение которых априорная и текущая
виды информации обрабатываются и взаимодействуют. Эти исследования позволяют
установить, на каком уровне нервной системы
и с участием каких механизмов происходит отбор значимой информации.
В настоящее время экспериментально показано, что в зависимости от задержки между моментами включения прайма и целевого стимула реализуются разные механизмы обработки зрительной информации и процессов произвольного селективного внимания [Черенкова, 2021].
В последнее время все чаще высказывается предположение, что
одной из причин особенностей восприятия у людей с РАС является именно
неспособность эффективно использовать априорную информацию [Soroor, 2022]. Сравнительный
анализ работ разных авторов свидетельствует об отсутствии единого мнения
о характеристиках проявления прайминг-эффекта при аутизме и РАС. Одни
исследователи указывают на отсутствие различий между людьми с аутизмом
и нейротипичными испытуемыми [Wilson, 2019]. Другие работы приводят данные об ином
характере проявления эффекта прайминга у аутистов [Soroor, 2022]. При этом люди
с аутизмом могут как превосходить испытуемых из контрольных групп по
показателям реакции (например, при выделении локальных элементов,
присутствующих в прайме и целевых стимулах) [Vanmarcke, 2017], так и испытывать сравнительно
бόльшие трудности, возникающие при выделении глобальных элементов объектов,
использовании в качестве прайма и целевого стимулов сложных изображений,
изображений неопределенной конфигурации [Pokhoday, 2019; Soroor, 2022; Van der Hallen, 2016].
В последние годы убедительно показано, что при аутизме в значительной мере страдают процессы, связанные с обработкой информации на нижних уровнях зрительной системы [Macaluso, 2016]. В связи с этим использование в процедуре прайминга объектов, содержащих простые признаки стимулов (такие как ориентация, пространственная частота), весьма актуальны для определения характеристик зрительного восприятия при аутизме.
Большинство рассмотренных исследований были проведены с
участием людей, имеющих аутистический спектр, без контроля их уровня интеллекта
и различий
в степени проявления тяжести РАС, тогда как работы, посвященные анализу
проявления прайминга при сниженном уровне интеллекта и разной выраженности РАС,
единичны [Черенкова, 2020а; McLaughlin, 2021]. При этом авторы концентрируют внимание на анализе прайминга у
лиц с РАС подросткового и зрелого возрастов [Vanmarcke, 2017]. Однако одним их критических
периодов развития процессов восприятия и произвольного внимания, которые
тестируются при прайминге, является дошкольный возраст. Изучение проявления
прайминга у детей данной возрастной группы позволит установить особенности и
этапы формирования зрительного восприятия при РАС.
Гипотеза исследования состояла в том, что у
детей дошкольного возраста
с РАС, имеющих разную степень выраженности расстройства, а также более низкий
уровень интеллекта и речевого развития по сравнению с нейротипичными
сверстниками, наблюдается измененный характер взаимодействия элементарных
признаков объектов (ориентация линий и их пространственная частота) на разных
временных этапах восприятия зрительной информации.
В задачи исследования входило сравнение характеристик и временной динамики зрительного прайминга при использовании простых зрительных стимулов у детей дошкольного возраста с типичным развитием и с разным уровнем выраженности РАС.
Методы исследования
Выборка. В исследовании приняли участие 20 детей с типичным развитием (из них 5 девочек, возраст — 6,1–6,9 лет) и 60 детей с РАС (из них 4 девочки, возраст — 6,1–7,0 лет). Все дети посещали дошкольное отделение начальной школы № 687 «Центр реабилитации ребенка» г. Санкт-Петербурга.
Дети с типичным развитием были протестированы с
помощью теста
Д. Векслера для детей дошкольного возраста (WHHSI), адаптированного в России
[Ильина, 2009]. Анализ результатов показал, что вербальный IQ варьировал от 110 до 128
баллов, невербальный IQ — от 116 до 134 баллов, что соответствовало уровню
нормативного развития (табл.).
Таблица
Характеристики обследуемых субвыборок испытуемых
|
Группа детей |
Количество детей |
Возраст |
Оценка тяжести РАС |
Оценка IQ |
Уровень общего недоразвития речи |
|
M±SD; диапазон |
M±SD; диапазон |
M±SD; диапазон в баллах |
|||
|
Дети |
20 (15 мальчиков, |
6,4±0,4; 6,1–6,9 |
|
По Векслеру: Вербальный IQ 119,0±9,0; 110,0–128,0 Невербальный IQ 125,0±9,0; 116,0–134,0 |
|
|
Дети |
20 (все мальчики) |
6,7±0,3; 6,3–7,1 |
32,0±2,0; 30,0–34,0 |
по LIPS Невербальный IQ 65,0±6,0; 59,0–70,0 |
ОНР II (формулируют предложения) |
|
Дети |
20 (18 мальчиков, |
6,6±0,4; 6,1–7,1 |
39,0±3,0; 36,0–42,0 |
по LIPS Невербальный IQ 52,0±4,0; 48,0–56,0 |
ОНР II-I (говорят отдельные слова) |
|
Дети |
20 (18 мальчиков, |
6,8±0,2; 6,5–7,2 |
47,0±3,0; 44,0–49,0 |
по LIPS Невербальный IQ 45,0±4,0; 41,0–49,0 |
ОНР I (произносят отдельные звуки) |
Отбор детей с РАС проводился консилиумом специалистов Центра реабилитации ребенка. Все дети в соответствии с МКБ-10 имели диагноз «РАС, обусловленное органическим заболеванием головного мозга, сопровождающемся задержкой психического развития, на резидуально-органической основе». Выраженность РАС устанавливалась по Оценочной шкале раннего детского аутизма — CАRS (Childhood Autism Rating Scale) [Schopler, 1980]. В соответствии с результатами диагностики по CARS были выделены три группы: I группа — дети с легкой формой РАС (30–34 балла), II группа — с умеренной тяжестью РАС (36–42 балла) и III группа — дети с тяжелой формой РАС (44–49 баллов).
Уровень невербального IQ у детей с РАС оценивали по Международной шкале продуктивности Лейтер — LIPS (Leiter International Performance Scale) [Roid, 2013]. У детей I группы показатели невербального IQ составили 59–70 баллов, у детей второй группы II группы — 48–56 баллов и у детей III группы — 44–49 баллов. Уровень речевого развития определялся c помощью анализа речевых карт [Нищева, 2007] и у детей I группы соответствовал ОНР II, у детей II группы — ОНР II-I, а у детей III группы — ОНР I (табл.).
Все испытуемые использовали правую руку в качестве ведущей и не имели проблем со зрением и слухом. Родители детей выразили добровольное согласие на их участие в исследовании. Проведение исследования одобрено Этическим комитетом Санкт-Петербургского государственного университета.
Стимулы. В качестве целевых стимулов
использовались изображения вертикальных и горизонтальных линий с частотой
следования 10 циклов на градус. В качестве прайм-стимулов в центре экрана
компьютера предъявлялась вертикальная решетка с частотой следования линий 2
цикла на градус из базы данных «Sutterstock». Яркость фона экрана компьютера
составляла 30 кд/м2, максимальная яркость изображения — 60
кд/м2, размер изображения — 8 угловых градусов, длительность
предъявления целевых стимулов составляла 200 мс,
а прайм-стимула — 100 мс. Межстимульный интервал (МСИ) между окончанием
предъявления прайма и началом предъявления тестового стимула варьировал от 50
до 600 мс (рис. 1).

Рис. 1. Схема предъявления стимулов
Процедура исследования. Тестирование
испытуемых проводилось в знакомой обстановке. Испытуемый удобно располагался
перед компьютером, расстояние от экрана монитора не превышало 50 см.
Освещенность помещения составляла 120 люкс. Для предъявления стимулов
использовали ноутбук Samsung R40-1 с размером экрана 17 дюймов с установленной
программой PsyTask v. 1.50.12. (ООО «Мицар»,
г. Санкт-Петербург, Россия).
Перед началом тестирования проводилось обучение задаче по различению двух целевых стимулов. В ответ на предъявления первого стимула необходимо было нажать на левую клавишу компьютера (20 попыток), а в ответ на предъявления второго — на правую клавишу компьютера (20 попыток). Все стимулы предъявлялись в случайном порядке с интервалом между пробами от 1,5 до 2,5 с.
При тестировании в равном количестве и в случайном порядке
предъявлялись комбинации целевых и прайм-стимулов. Было проведено два теста, в
каждом из которых применялись МСИ четырех значений (50, 100, 150 и 200 мс; 300,
400, 500
и 600 мс). В каждом тесте предъявлялось 80 сочетаний стимулов (по 20 проб
с каждым значением МСИ) с интервалом между пробами в 1,5–2,5 с. Время реакции
(ВР) регистрировалось с помощью программы PsyTask во время обучения
и тестирования.
Анализ результатов. Статистическая обработка
данных проводилась
в программе Statistica v. 13. Распределение переменных по всей выборке
соответствовало нормальному (тест Колмогорова–Смирнова: d=0,25; р=0,200), что
позволило использовать для анализа параметрические критерии — t-тест
Стьюдента
и дисперсионный анализ ANOVA для повторных измерений. Для сравнения изменения
параметров реакций в зависимости от условий предъявления стимулов (с праймом
или без, предъявление прайма и тестового стимулов с разными МСИ) использовали
модель анализа для внутригрупповых зависимых факторов. При статистическом
анализе зависимости величины прайминг-эффекта от выраженности РАС и уровня
интеллекта факторы «группа» (группа детей в соответствии с уровнем выраженности
РАС) и «IQ» (показатель интеллекта) рассматривали в качестве независимых. Для
анализа ВР отбирались только правильные реакции (92,66% из всех проб). Значения
ВР, которые превышали 2 стандартных отклонения от средней величины, не
включались в обработку данных (2,37% из всех проб). При статистической
обработке для каждого испытуемого определялась величина прайминг-эффекта при
каждом значении МСИ по разнице между средними значениями ВР на предъявление
только целевого стимула и сочетания целевого
и прайм-стимулов. Далее полученные разности усреднялись по группам испытуемых.
Достоверность различий данных устанавливалась с помощью метода парных сравнений
Стьюдента. Достоверными считали различия при р≤0,05.
Результаты
Обучение. Дети с нейротипичным
развитием и дети с легкой формой РАС
в первый день тренировки верно дифференцировали целевые стимулы в 94–98%
(М=96,4; SD=0,2) случаев. У остальных детей процесс обучения занимал три дня.
Число правильных ответов к концу обучения варьировало в пределах 86–94%
(М=90,6; SD=0,5). ВР на линии разной ориентации сильно варьировало в пределах
выборки. При этом ВР у детей с типичным развитием и легкой формой РАС
достоверно не отличалось (M=1003 мс, SD=95, диапазон — 849–1099 мс и М=1052 мс,
SD=105, диапазон — 788–1184 мс соответственно, t=1,22, р=0,240). В то же
время
у детей с умеренной и тяжелой формами РАС ВР было достоверно больше по
сравнению с предыдущими группами (М=1734 мс, SD=233, диапазон — 1418–1890 мс и
М=1746 мс, SD=171, диапазон — 144–1860 мс соответственно, t=5,41, p˂0,001).
Между значениями ВР у групп детей с умеренной и тяжелой формой РАС достоверных
различий выявлено не было (t=0,74, р=0,670).
Тестирование нейротипичных детей. Предъявление
вертикальных линий
с низкой частотой следования приводило к достоверному снижению ВР при
идентификации вертикальных линий с высокой частотой следования (M=1001 мс,
SD=93, диапазон — 849–1080 мс при обучении и M=902 мс, SD=43, диапазон —
885–931 мс при тестировании; t=4,65, p˂0,001). В случае идентификации
горизонтальных линий с высокой частотой следования значимых изменений ВР не
наблюдалось (M=998 мс, SD=67, диапазон — 849–1096 мс при обучении и M=1004 мс,
SD=69, диапазон — 978–1023 мс при тестировании; t=0,95, p=0,450) (рис. 2).
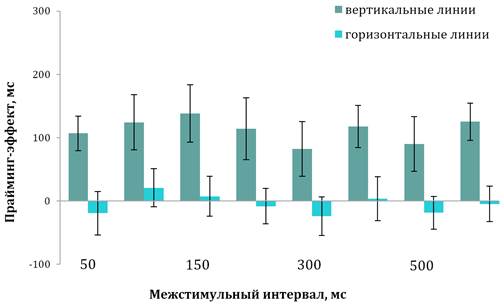
Рис. 2. Временная динамика прайминг-эффекта у детей с типичным развитием
Примечания. Вертикальные линии обозначают доверительный интервал при р≤0,05.
Дисперсионный анализ показал зависимость величины прайминг-эффекта от типа идентифицируемого (целевого) стимула (F(1; 158)=170,69, p˂0,001). Не было отмечено зависимости этой величины от IQ (F(1; 158)=7,69, p=0,350). В то же время зависимость величины прайминг-эффекта от МСИ не была значимой ни для реакций на вертикальные линии (F(7; 152)=0,76, p=0,620), ни для реакций на горизонтальные линии (F(7; 152)=0,91, р=0,510). Однако более детальный анализ продемонстрировал, что при идентификации вертикальных линий при МСИ от 50 до 150 мс величина прайминг-эффекта прогрессивно растет (F(2; 57)=19,75, p˂0,001), а при МСИ от 200 до 600 мс остается стабильной (F(4; 95)=1,58, p=0,090).
Тестирование детей с легкой формой РАС. У детей этой группы при введении прайм-стимула наблюдалась иная картина (рис. 3). ВР снижалось не только при предъявлении вертикальных линий (М=1001 мс, SD=105, диапазон — 927–1139 мс при обучении и M=961 мс, SD=109, диапазон — 874–1062 мс при тестировании; t=2,07, p=0,030). Идентификация горизонтальных линий также сопровождалась уменьшением ВР (M=1068 мс, SD=201, диапазон — 788–1184мс при обучении и M=995 мс, SD=114, диапазон — 922–1074 мс при тестировании; t=2,14, p=0,020). Попарное сравнение ВР на экспозицию вертикальных и горизонтальных линий после прайма не выявило достоверных различий ни при одном значении МСИ (t=2,09, p=0,150).
Дисперсионный анализ данных показал, что величина прайминг-эффекта значимо зависит от МСИ (F(7; 152)=14,91, p˂0,001). Однако детальный анализ показал, что при МСИ в 50, 100 и 150 мс снижение ВР на оба типа целевых стимулов не было значимым (t=1,17, p=0,250; t=0,29, p=0,370 и t=0,92, p=0,360, соответственно). Между тем в интервале задержек от 200 до 600 мс уменьшение ВР было достоверным и стабильным независимо от значения МСИ (F(4; 95)=31,68, p˂0,001). Зависимость величины прайминг-эффекта от уровня интеллекта не была значимой (F(1; 152)=170,69, p˂0,001).
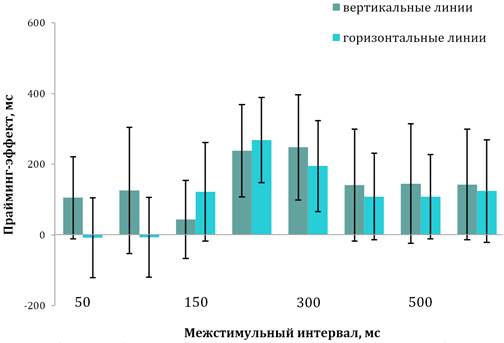
Рис. 3. Временная динамика прайминг-эффекта у детей с легкой формой РАС
Примечания. Вертикальные линии обозначают доверительный интервал при р≤0,05.
Тестирование детей с умеренной формой РАС. Как
и у детей с легкой формой РАС, при введении прайм-стимула наблюдалось снижение
ВР на предъявление как вертикальных (М=1739 мс, SD=234; диапазон — 1524–1900 мс
при обучении
и M=1525 мс; SD=187; диапазон — 1407–1716 мс при тестировании; t=2,48,
p=0,010), так и горизонтальных линий (M=1628 мс, SD=235, диапазон — 1523–1723
мс при обучении и M=1632 мс, SD=214, диапазон — 1461–1735 мс при тестировании;
t=2,57, p=0,010). ANOVA не выявил зависимости прайминг-эффекта от типа целевого
стимула (F(1; 158)=2,97, р=0,540) (рис. 4).
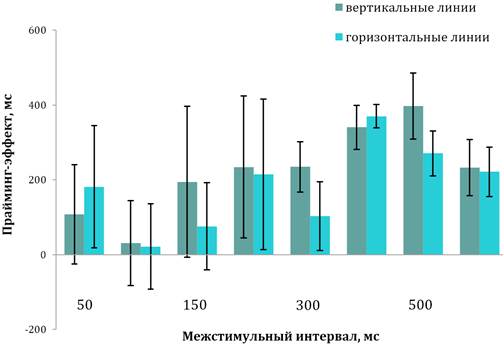
Рис. 4. Временная динамика прайминг-эффекта у детей с умеренной формой РАС
Примечания. Вертикальные линии обозначают доверительный интервал при р≤0,05.
Как и у детей с легкой формой РАС, анализ динамики изменения
прайминг-эффекта показал значимую зависимость от увеличения МСИ при
идентификации
и горизонтальных, и вертикальных линий (F(7; 152)=12,27, p˂0,001), но не выявил
зависимости прайминг-эффекта от величины IQ (F(2; 158)=2,47, p=0,150).
Анализ изменения прайминг-эффекта при разных значениях МСИ установил, что при МСИ в интервале от 50 до 150 мс величина прайминг-эффекта не достигала значимых значений (F(2; 57)=2,18, p=0,060). При МСИ от 200 до 600 мс ВР изменялось в разном направлении. При МСИ 200 и 300 мс значение прайминг-эффекта было значимым на оба стимула (t=4,41, p=0,040), при задержках 400 и 500 мс достоверно возрастало (t=9,48, p=0,010), а при 600 мс достоверно снижалось (t=8,41, p=0,020) (рис. 4).
Тестирование детей с тяжелой формой РАС. Как и
у детей с легкой
и умеренной формами РАС, у детей данной группы при введении прайм-стимула
происходило снижение ВР как в случае экспозиции вертикальных (М=1747 мс,
SD=138, диапазон — 1592–1951 мс при обучении и M=1603 мс, SD=199, диапазон —
1491–1614 мс при тестировании; t=2,31, р=0,030), так и горизонтальных линий
(M=1745 мс, SD=193, диапазон — 1441–1967 мс при обучении и M=1632 мс, SD=214,
диапазон — 1461–1735 мс при тестировании; t=2,54, р=0,020) (рис. 5).
Дисперсионный анализ не выявил значимой зависимости величины
прайминг-эффекта от типа стимула (F(1; 158)=2,09, p=0,150) и от величины IQ
(F(1; 158)=2,24, р=0,180). Анализ временной динамики прайминг-эффекта показал
значимую зависимость величины прайминг-эффекта от МСИ при идентификации
и горизонтальных, и вертикальных линий, так же как у предыдущих групп детей
(F(7; 152)=2,69, p=0,010). Анализ изменения прайминг-эффекта при разных
значениях МСИ установил, что при МСИ в интервале от 50 до 150 мс величина
прайминг-эффекта не достигала значимых значений (F(2; 58)=2,18, p=0,060). Было
установлено, что, как и у детей с умеренной формой РАС, при МСИ в 200 и 300 мс
значение прайминг-эффекта на оба стимула достоверно возрастало (t=7,89,
p=0,010), а при МСИ в 400, 500 и 600 мс вновь не достигала значимых значений
(t=1,41, p=0,080) (рис. 5). Сравнение средней величины прайминг-эффекта у
разных групп детей не выявил ее значимой зависимости от факторов «группа»
(F(3;76)=1,17, p=0,210) и «IQ» (F(1;76)=2,19, p=0,340). Детальный анализ
проявления прайминга при разных МСИ позволил выявить достоверность зависимости
прайминг-эффекта от данных факторов только в интервале задержек 50–150 мс
(F(3;76)=68,87, p˂0,001; F(1;76)=59,47, p˂0,001).
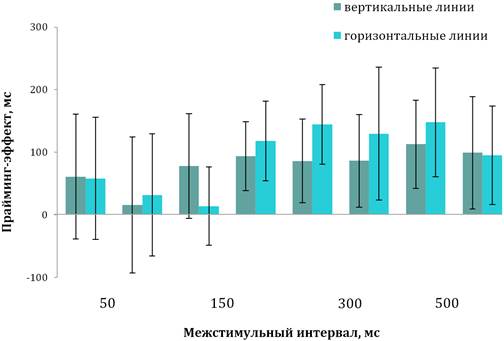
Рис. 5. Временная динамика прайминг-эффекта у детей с тяжелой формой РАС
Примечания. Вертикальные линии обозначают доверительный интервал при р≤0,05.
Полученные результаты показали, что у детей с РАС в отличие
от нейротипичных детей наблюдается измененный характер зрительного прайминга
при использовании простых зрительных стимулов. Скорость реакции возрастает на
линии разной ориентации независимо от отсутствия/присутствия этой ориентации в
прайме. У детей с разной выраженностью РАС наблюдаемый эффект уменьшения
времени реакции отмечается при разных временных задержках между праймом
и целевым стимулом.
Обсуждение результатов
Исследование зрительного прайминга у детей дошкольного возраста с РАС разной тяжести позволило выявить особенности временной динамики эффекта прайминга при идентификации простых зрительных стимулов.
В первую очередь необходимо отметить, что у всех детей с РАС влияние предъявления прайма не зависит от свойств идентифицируемого изображения. Если для нейротипичных детей характерно уменьшение ВР только на тот стимул, который включает ориентацию линий, аналогичную ориентации линий в прайме, то у детей с РАС ускорение реакции возможно и на стимул, включающий иную ориентацию линий. Наличие четкой диссоциации влияния прайма у нейротипичных детей свидетельствует о том, что в возрасте шести лет уже сформированы механизмы выделения общего для прайма и целевого стимула ведущего признака, что подтверждают работы ряда авторов [Черенкова, 2021; Kristjansson, 2019]. У детей с РАС проявление этих механизмов не наблюдается, что согласуется с работами, которые показали отсутствие способности непроизвольно концентрировать внимание на глобальных элементах объектов внешнего мира при РАС [4–6]. Встает вопрос о процессах обработки информации, которые лежат в основе специфической реакции, выявленной у детей с РАС. Ответить на этот вопрос помогает анализ временной динамики прайминга.
У нейротипичных детей достоверное влияние прайма отмечается уже при МСИ в 50 мс. Нейрофизиологические данные свидетельствуют о том, что в период задержек целевого стимула от начала прайма (SOA) до 300 мс нейроны ассоциативных областей коры у обезьян кодируют факт наличия двух стимулов на входе сенсорной системы [Carlson, 2013]. В нашей модели прайминга при МСИ, равном 50 мс, SOA составляет 150 мс. Следовательно, уже в первые 150 мс дети с типичным развитием способны выделить значимый элемент стимула, являющийся общим для прайма и целевого объекта.
У всех обследованных детей с РАС при МСИ до 150 мс (SOA=250
мс) достоверное влияние прайма на идентификацию целевых стимулов отсутствовало.
Данные литературы о временных параметрах взаимодействия сенсорных входов
позволяют выдвинуть несколько причин, обусловливающих полученные результаты. С
одной стороны, была предложена модель независимого взаимодействия сенсорных
входов, которая характерна для ранних периодов онтогенеза [Miller, 1982]. Согласно этой
модели, каждый сенсорный вход может самостоятельно конкурировать за выход на
соответствующую реакцию организма. В результате можно наблюдать
последовательную череду реакций то на один, то на другой сигнал. Согласно этой
точке зрения, отсутствие влияния прайма у детей с РАС может говорить
о незрелости процессов взаимодействия сенсорной информации. Аналогичный тип
взаимодействия проявляется у нейротипичных детей четырех лет при использовании
процедуры прайминга с гетеросенсорными объектами [Черенкова, 2020; Черенкова, 2020а].
С другой стороны, имеются данные о задержке процесса
взаимодействия сенсорных входов на уровне зрительных областей коры при РАС. В
отношении стимулов с разной пространственной частотой показано, что при РАС
информация по низкочастотному каналу приходит значительно позже по
сравнению
с информацией по высокочастотному каналу [Sutherland, 2010]. Это означает, что процесс
взаимодействия зрительных сигналов может быть задержан по времени от начала
действия прайма — в нашем случае на 250 мс.
Интервал между праймом и целевым стимулом от 300 до 600 мс
рассматривается в нейрофизиологических исследованиях и работах по праймингу как
время, необходимое для сличения уже опознанных объектов и сдвига внимания в
сторону идентифицируемых стимулов, требующих соответствующей реакции [Carlson, 2013]. В
этот период влияние прайма постепенно нивелируется, что отражается
в отсутствии прироста эффекта облегчения реакции. Это хорошо просматривается у
нейротипичных детей. Если при задержках до 150 мс отмечается сокращение ВР, то
после 200 мс его зависимость от МСИ достоверно не выражена. При РАС, напротив,
именно в этот период эффект прайминга приобретает достоверные значения. Дети
быстрее реагируют на предъявление как вертикальных, так и горизонтальных линий.
Подобное проявление прайминга может свидетельствовать о нарушении процессов
нисходящей регуляции переключения внимания между опознанными стимулами, что
было показано в исследовании, проведенном на взрослых
и подростках с аутизмом [Macaluso, 2016]. При этом можно предположить, что у детей с РАС
длительный процесс взаимодействия информации о прайме и целевом стимуле влечет
за собой позднее включение механизмов переключения внимания от прайма к
целевому стимулу. Следовательно, отсутствие сдвига внимания в сторону целевых
стимулов может быть связано как с изменением механизмов взаимодействия
информации на уровне проекционных областей коры (первые 250 мс), так и от
характера включающейся позднее регуляции этого процесса со стороны зрительных
ассоциативных областей коры [Sutherland, 2010].
У детей с тяжелой формой РАС достоверный прайминг-эффект отмечен только при МСИ в 200 и 300 мс. В отличие от детей первых двух групп, в интервале МСИ от 400 до 600 мс прайминг-эффект проявляется случайно. Можно предположить, что процесс сличения уже опознанных стимулов в этом случае происходит на независимой конкурентной основе, как в модели независимого взаимодействия входной информации, а реакция реализуется в ответ то на один, то на другой целевые стимулы. Однако после опознания стимулов, использованных в процедуре прайминга, такое независимое соперничество взаимодействующей информации возможно только вследствие изменения механизма нисходящей регуляции процессов внимания [Sutherland, 2010].
Заключение
Полученные в ходе исследования результаты свидетельствуют о
специфических особенностях проявления зрительного прайминга у детей дошкольного
возраста
с РАС по сравнению с детьми с типичным развитием. У нейротипичных детей прайм
вызывает уменьшение ВР только на один целевой стимул. У детей с РАС влияние
априорной информации отсутствует при задержке целевых стимулов относительно
прайма от 50 до 150 мс. У детей с легкой и умеренной формами РАС уменьшение ВР
наблюдается для всех целевых стимулов в интервале МСИ от 200 до 600 мс, а у
детей с тяжелой формой РАС — при МСИ в 200 и 300 мс. Наблюдаемая временная
динамика взаимодействия зрительной информации может свидетельствовать об
изменении процессов мономодального взаимодействия на нижних уровнях сенсорной
системы и нарушений сдвига внимания между праймом и целевым стимулом,
обусловленным как в восходящей системе регуляции внимания, так
и в системе нисходящей регуляции внимания в зависимости от уровня проявления
аутистических симптомов.
К ограничениям проведенного исследования необходимо отнести
сравнение групп детей с РАС только с нейротипичными сверстниками. Для того,
чтобы определить, насколько наблюдаемые изменения зрительного прайминга
характерны только для РАС, необходимо провести дополнительное обследования
детей с задержкой психического развития, имеющих схожее снижение
интеллектуального и речевого развития. Группы детей с РАС также были выравнены
не только по выраженности аутистических симптомов, но и по уровню невербального
интеллекта и нарушений речевого развития. Это условие не позволяет установить
зависимость зрительного прайминга от каждой из этих переменных. Решение этого
вопроса требует дополнительных обследований детей
с одинаковой выраженностью РАС, но значительно отличающихся по уровню
интеллектуального и речевого развития.
Литература
- Ильина М.Н. Психологическая оценка интеллекта у детей. СПб: Питер, 2009. 366 с.
- Нищева Н.В. Речевая карта ребенка. М.: Детство-Пресс, 2007. 265 с.
- Черенкова Л.В., Соколова Л.В. Возрастная динамика процессов зрительного прайминга // Российский физиологический журнал. 2021. Том 107. № 9. С. 1112–1125. DOI: 10.31857/S086981392109003X
- Черенкова Л.В., Соколова Л.В. Исследование процессов антиципации у детей дошкольного возраста с атипичным развитием // Психологический журнал. 2020. Том 41. № 3. С. 66–77. DOI: 10.31857/S020595920008518-5
- Черенкова Л.В., Соколова Л.В. Проявление прайминг-эффекта у детей дошкольного возраста с расстройством аутического спектра // Физиология человека. 2020. Том 46. № 2. С. 38–46. DOI: 10.31857/S0131164620010051
- Amso D., Haas S., Tenenbaum E. et al. Bottom-up attention orienting in young children with autism // Journal of Autism and Developmental Disorders. 2014. Vol. 44. № 2. P. 664–673. DOI: 10.1007/s10803-013-1925-5
- Carlson T., Tovar D.A., Alink A. et al. Representational dynamics of object vision: the first 1000 ms // Vision. 2013. Vol. 13. № 1. P. 1–19. DOI: 10.1167/13.10.1
- Haigh S.M. Variable sensory perception in autism // Europe Journal of Neuroscience. 2018. Vol. 47. № 6. P. 488–766. DOI:10.1111/ejn.1360
- Happe F., Frith U. The weak central coherence account: detail-focused cognitive style in autism spectrum disorders // Journal of Autism and Developmental Disorders. 2006. Vol. 36. № 1. P. 5–25. DOI: 10.1007/s10803-005-0039-0
- Janiszewski C., Wyer R.S. Content and process priming: A review // Journal of Consumer Psychology. 2014. Vol. 24. № 1. P. 96 – 118. DOI: 10.1016/j.jcps.2013.05.006
- Jurkat S., Gruber M., Kärtner J. The effect of verbal priming of visual attention styles in 4- to 9-year-old children // Cognition. 2021. Vol. 212. 3. P. 104–181. DOI: 10.1016/j.cognition.2021.104681
- Kimchi R. The perception of hierarchical structure: Oxford Handbook of Perceptual Organization / J. Wagemans (ed.). Oxford, UK: Oxford University Press, 2015. P. 129–149. DOI: 10.1177/1747021818766848
- Kristjansson A., Asgeirsson A.G. Attentional priming: Recent insights and current controversies // Current Opinion in Psychology. 2019. Vol. 29. № 1. P. 71–75. DOI: 10.1016/j.copsyc.2018.11.0133
- Macaluso E., Noppeney U., Talsma D. et al. The curious incident of attention in multisensory integration: Bottom-up vs. top-down // Multisensory Research. 2016. Vol. 29. № 6–7. P. 557–583. DOI: 10.1163/22134808-00002528l
-
McLaughlin C.S., Grosman H., Grosman S. et al.
Reduced engagement of visual attention in children with autism spectrum
disorder // Autism. 2021. Vol. 25. № 2.
P. 1362–1392. DOI: 10.1177/13623613211010072 - Miller J. Divided attention: Evidence for coactivation with redundant signals // Cognitive Psychology. 1982. Vol. 14. № 2. P. 247–279. DOI: 10.1016/0010-0285(82)90010-X
- Mo S., Liang L., Bardikoff N. et al. Shifting visual attention to social and non-social stimuli in Autism Spectrum Disorders // Research in Autism Spectrum Disorders. 2019. Vol. 65. № 1. P. 56–64. DOI: 10.1016/j.rasd.2019.05.006
- Pokhoday M., Shtyrov Y., Myachykov A. Effects of visual priming and event orientation on word order choice in Russian sentence production // Frontiers in Psychology. 2019. Vol. 10. Article 1661. DOI: 10.3389/fpsyg.2019.01661
- Robertson C.E., Baron-Cohen S. Sensory perception in autism // Nature Review Neuroscience. 2017. Vol. 18. № 11. P. 671–684. DOI: 10.1038/nrn.2017.112
- Roid G.H., Mille L.J., Pomplun M. et al. Leiter‑3: Leiter International Performance Scale: Manual / G. Roid (ed.). Wood Dale, Illinois: Stoelting Co., 2013. P. 5–307.
- Schopler E., Reichler R.J., DeVellis R.F. et al. Toward objective classification of childhood autism: Childhood Autism Rating Scale (CARS) // Journal of Autism and Developmental Disorders. 1980. Vol. 10. № 1. P. 91–103. DOI: 10.1007/BF02408436
- Shomstein S., Kravit D.J., Behrmann M. Attentional control: temporal relationships within the fronto-parieatal network // Neuropsychoiogia. 2012. Vol. 50. № 6. P. 1202–1210. DOI: 10.1016/j.neuropsychologia.2012.02.009
- Soroor G., Mokhtari S. Pouretemad H. Priming global processing strategy improves the perceptual performance of children with Autism Spectrum Disorders // Journal of Autism and Developmental Disorders. 2022. Vol. 52. № 4. P. 1019–1029. DOI:10.1007/ s10803-021-05007-7
- Sutherland A., Crewther D.P. Magnocellular visual evoked potential delay with high autism spectrum quotient yields a neural mechanism for altered perception // Brain. 2010. Vol. 133. № 4. P. 2089–2097. DOI: 10.1093/brain/awq122
- Van der Hallen R., Evers K., Boet B. et al. Visual search in ASD: Instructed versus spontaneous local and global processing // Journal of Autism and Developmental Disorders. 2016. Vol. 46. № 5. P. 3023–3036. DOI: 10.1007/s10803-016-2826-1
- Vanmarcke S., Noens I., Steyaert J. et al. Spatial frequency priming of scene perception in adolescents with and without ASD // Journal of Autism and Developmental Disorders. 2017. Vol. 47. № 2. P. 2023–2038 DOI:10.1007/s10803-017-3123-3
- Wilson C.E., Saldaña D. No evidence of atypical attentional disengagement in autism: A study across the spectrum // Autism: The International Journal of Research and Practice. 2019. Vol. 23. № 3. P. 677–688. DOI: 10.1177/2F1362361318768025
Информация об авторах
Метрики
Просмотров web
За все время: 926
В прошлом месяце: 24
В текущем месяце: 13
Скачиваний PDF
За все время: 285
В прошлом месяце: 8
В текущем месяце: 4
Всего
За все время: 1211
В прошлом месяце: 32
В текущем месяце: 17