Экспериментальная психология
2022. Том 15. № 1. С. 56–71
doi:10.17759/exppsy.2022150104
ISSN: 2072-7593 / 2311-7036 (online)
Изменение ЭЭГ-ритмов и вариабельности ритма сердца в ответ на холод в зависимости от параметров произвольного внимания у молодых людей
Аннотация
Общая информация
Ключевые слова: ЭЭГ, вариабельность сердечного ритма, произвольное внимание, холод
Рубрика издания: Психофизиология
Тип материала: научная статья
DOI: https://doi.org/10.17759/exppsy.2022150104
Финансирование. Работа выполнена по программе ФНИР ФГБУН ФИЦКИА УрО РАН, № AAAA-A19-119120990083-9.
Благодарности. Авторы выражают благодарность зав. лаборатории регуляторных механизмов иммунитета кандидату биологических наук А.В. Самодовой за возможность использования в исследовании холодовой камеры и за помощь в сборе первичного материала сотруднику лаборатории биоритмологии ФИЦКИА УрО РАН доктору медицинских наук Д.Б. Дёмину.
Получена: 14.01.2021
Принята в печать:
Для цитаты: Кривоногова Е.В., Кривоногова О.В., Поскотинова Л.В. Изменение ЭЭГ-ритмов и вариабельности ритма сердца в ответ на холод в зависимости от параметров произвольного внимания у молодых людей // Экспериментальная психология. 2022. Том 15. № 1. С. 56–71. DOI: 10.17759/exppsy.2022150104
Полный текст
Введение
В настоящее время в связи со значимостью развития территорий Арктической зоны оценка адаптационных возможностей человека и выявление интегративных показателей, отражающих эффективность приспособительных реакций организма, остается одним из актуальных направлений общей психологии и психологии труда. Адаптация к дискомфортным факторам среды Севера (низкие температуры, низкая солнечная активность) обеспечивается системными реакциями организма и выражается в его непрерывном приспособлении к условиям природной среды и одновременно к трудовой и социальной деятельности в данных условиях [1; 5]. В условиях Севера происходит напряжение адаптационных механизмов человека вследствие необходимости формирования приспособительных реакций для поддержания гомеостаза [Максимов, 2021]. В ряде исследований показано замедление функции переключения внимания и удлинение латентного времени Р300 у молодежи, проживающей на Севере, в сравнении с ровесниками, проживающими в южных регионах [Кривоногова, 2020; Солонин, 2019]. Взаимодействие между вегетативной нервной системой и нейронными сетями более высокого порядка имеет решающее значение для когнитивной регуляции. Способность удерживать и переключать внимание — важный компонент саморегуляции и приспособления организма [Posner, 2020]. Адаптационные реакции индивидуальны и реализуются у разных лиц с различной степенью участия функциональных систем [Марков, 2018]. О характере приспособительных реакций можно судить по динамике отдельных показателей физиологических систем организма под воздействием функциональных нагрузок [Максимов, 2021]. Однако вопрос о механизмах взаимодействия вегетативной и центральной нервной системы, а также симпатической и парасимпатической нервной системы с точки зрения индивидуальных различий в приспособительных реакциях и обеспечения когнитивной деятельности в условиях адаптации человека к аномальным условиями жизнедеятельности остается малоизученным.
Цель работы состояла в оценке взаимосвязи индивидуально-типологических реакций биоэлектрической активности мозга и вариабельности ритма сердца и параметров произвольного внимания в условиях воздействия низких температур на организм человека.
Гипотезой исследования являлось предположение о том, что различия в показателях произвольного внимания, отражающих системную организацию когнитивных процессов, которые протекают на центральном и периферическом уровнях, определяются особенностями перестройки баланса вегетативной нервной системы, биоэлектрической активности мозга для поддержания температурного гомеостаза при воздействии холода.
Материалы и методы
В исследовании приняли участие 28 испытуемых мужского пола в возрасте 18—21 года (средний возраст 19,9±1,2 лет, практически здоровые, жители г. Архангельска). Исследование было одобрено Этическим комитетом Федерального исследовательского центра комплексного изучения Арктики имени академика Н.П. Лаверова УрО РАН (протокол № 2 от 28.03.2018). Все лица, участвующие в исследовании, подписали информированное согласие. Исследования проводились в зимний период года (январь—февраль) в первой половине дня. Индекс массы тела обследуемых не выходил за границы нормальных значений (от 18,5 кг/м2 до 24,9 кг/м2). Все обследуемые были в однотипной одежде — кроссовки, хлопчатобумажные брюки, футболка и медицинский халат.
Исследование воздействия холода на организм человека состояло из 3 этапов. 1-й этап: осуществлялась оценка параметров синхронной записи электроэнцефалограммы (ЭЭГ) и вариабельности сердечного ритма (ВСР) (5 минут), температуры кожного покрова кисти (Тк) и в слуховом проходе (Тсл) в теплом помещении (этап — фон). 2-й этап: с пятой по десятую минуты нахождения в холодовой камере «УШЗ-25Н» (-200С) производилась регистрация параметров ЭЭГ, ВСР, на пятой и десятой минуте пребывания в холоде — Тк и Тсл (этап — холод). 3-й этап: через 5 минут после выхода из холодовой камеры регистрировали ЭЭГ, ВСР (5 мин), Тк, Тсл (этап — тепло через 5 мин). Перед экспериментальным исследованием проводилась оценка произвольного внимания по тесту Тулуз—Пьерона в теплом помещении на основании параметров скорости (V), точности (К) выполнения заданий теста и количества ошибок (F). Нормативные значения и уровень произвольного внимания определялись по таблицам, представленным в методическом руководстве Л.А. Ясюковой [Ясюкова, 1997]. Скорость выполнения теста (знаков/мин) ниже 49 соответствовала низкой, в интервале от 50 до 62 — средней, от 63 до 77 — хорошей, выше 78 — высокой. Точность выполнения теста (коэффициент точности) ниже 0,91 соответствовала низкому уровню, в интервале от 0,92 до 0,95 — среднему, от 0,96 до 0,97 — средневысокому, выше 0,98 — высокому уровню.
Оценка ЭЭГ осуществлялась по спектральной мощности (СМ): α (8—13 Гц), β1 (14—24Гц), θ (4—7 Гц) — активности. ЭЭГ регистрировали по 16 каналам (Fp1, Fp2, F3, F4, F7, F8, C3, C4, P3, P4, T3, T4, T5, T6, O1, O2) по международной системе «10—20%» размещения электродов на поверхности головы (референтные электроды на мочках ушей) в состоянии спокойного бодрствования с закрытыми глазами в положении сидя с помощью портативного электроэнцефалографа (Нейрон-Спектр-СМ «Нейрософт», г. Иваново). Оценка состояния вегетативной нервной системы осуществлялась по показателям ВСР, которые оценивались с помощью АПК «Варикард» (г. Рязань). Использовали временной и спектральный виды анализа: частота сердечных сокращений (ЧСС, уд. в мин.), среднее квадратичное отклонение (SDNN, мс), отражающее суммарное значений показателей вегетативной регуляции кровообращения; квадратный корень из суммы квадратов разности величин последовательных пар RR-интервалов (RMSSD, мс) и процент количества пар последовательных кардиоинтервалов в кардиограмме, отличающихся более чем на 50 мс (PNN50%), отражают активность парасимпатического звена вегетативной регуляции, индекс напряжения регуляторных систем (SI, усл. ед.), мощность спектра ВСР в диапазоне высоких частот (HF, 0,4—0,15 Гц), в низкочастотном (LF, 0,04 до 0,15 Гц) и в сверхнизкочастотном диапазоне (VLF, 0,04—0,015 Гц); суммарная мощность спектра ВСР (ТР, мс2). Тк и Тсл регистрировали медицинским электронным инфракрасным термометром B.Well WF — 1000 (Швейцария). Статистическую обработку данных проводили в программе Statistica 6.0. Проверка распределения количественных данных на нормальность осуществлялась с использованием статистических критериев Shapiro—Wilk. Показатели ВСР, ЭЭГ, параметры произвольного внимания, Тк, Тсл представлены в виде медианы и 25- и 75-перцентилей (Ме (25р; 75р)). Для выделения однородных групп по СМ α- и β1-активности ЭЭГ в лобных отделах (F3, F4), параметрам ВСР (ТР, SI), показателям произвольного внимания (V, K) и температуры поверхности тела (Тсл, Тк) в фоне использовали кластерный анализ методом к-средних. Для проведения кластерного анализа было проведено z-преобразование данных с целью уменьшения асимметрии при распределении переменных.
Сравнения количественных данных для трех независимых групп проводили с помощью критерия Краскела—Уоллиса, двух независимых групп — критерия Манна—Уитни. Для анализа повторных измерений применяли критерий хи-квадрат Фридмана, при попарном сравнительном анализе — метод Вилкоксона, использовали критический уровень значимости менее 0,05 при сравнении с фоновыми значениями.
Результаты исследования
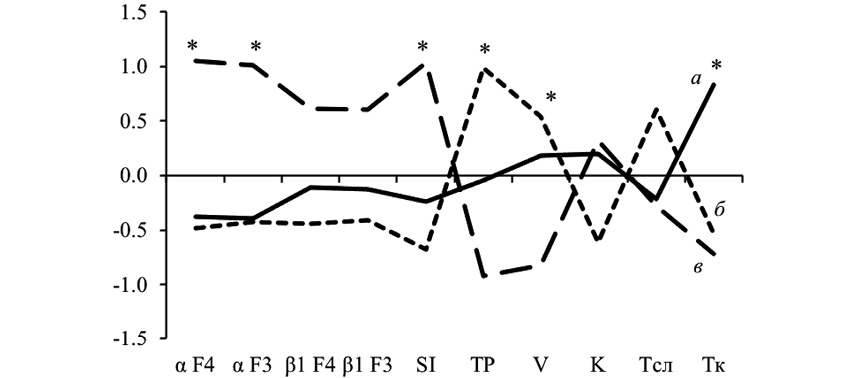
Рис. 1. Результаты кластерного анализа: а — группа I; б — группа II; в — группа III; αF4, αF3 —спектральная мощность α-активности ЭЭГ; β1F4, β1F3 — спектральная мощность β1-активности ЭЭГ; SI, TP — параметры ВСР; V— скорость; K — точность выполнения теста; * — p<0,01 уровень статистической значимости при сравнении между группами
Путем кластеризации было выделено три однородные группы испытуемых, различающиеся по параметрам произвольного внимания, биоэлектрической активности головного мозга и параметрам ВСР при фоновой оценке (рис. 1).
Анализ параметров произвольного внимания выявил, что у испытуемых групп I и II наблюдалась средняя скорость выполнения заданий теста, а у испытуемых группы III выявлена низкая скорость. По точности выполнения теста статистических значимых различий не выявлено между испытуемыми трех групп. Однако в группе I отмечался высокий уровень точности выполнения заданий, в то время как у испытуемых группы II было выявлено большее количество ошибок в тесте (табл. 1).
Таблица 1
Параметры произвольного внимания на основании оценки
показателей
всех испытуемых, Ме (25; 75)
|
Показатель |
Группа I N=12 |
Группа II N=8 |
Группа III N=8 |
p, Краскела—Уоллиса |
|
Скорость (V), знаков/мин. |
58,8(50,5;61,0) |
59,4(54,8;66,2) |
44,7(38,4;53,2)**II-III |
0,034 |
|
Количество ошибок (F) |
1,6(1,0;2,7) |
2,4(2,1;4,1)**II-III,I |
1,2(1,0;1,7) |
0,018 |
|
Точность (K) |
0,98(0,95;0,98) |
0,96(0,93;0,97) |
0,97(0,96;0,98) |
0,151 |
Примечание: «*» — р <0,05; «**» — p <0,01 уровень статистической значимости между группами (критерий Манна—Уитни).
Анализ температуры кожного покрова кисти рук (Тк) выявил различия между группами на этапе фоновой оценки (рис. 2) — у испытуемых групп II и III показатели Тк являются статистически более низкими, чем показатели Тк у испытуемых группы I. При воздействии холода обнаруживалась динамика снижения показателей — Тк снижалась во всех группах испытуемых, при этом в группе III показатели Тк обнаруживали более низкие значения по сравнению с показателями испытуемых группы I. Оценка аналогичных показателей через 5 минут после пребывания на холоде обнаруживает отсутствие восстановления Тк у испытуемых.

Рис. 2. Показатели температуры поверхности кожи кисти у испытуемых на различных этапах исследования: а — группа I; б — группа II; в — группа III; Δ — p<0,01 — при сравнении между этапами исследования (критерий Вилкосона); ** — p<0,01 — между группами (критерий Манна—Уитни)
Анализ полученных данных свидетельствует об отсутствии существенных различий в показателях температуры в слуховом проходе у всех групп испытуемых (рис. 3), однако указывают на снижение их значений после 5-минутного воздействия холода по сравнению со значениями показателей при фоновой оценке. Через 10 минут пребывания на холоде отмечалось существенное снижение показателей Тсл во всех группах, при этом в группе II наблюдалась более слабая динамика снижения показателей Тсл по сравнению с другими группами.

Рис. 3. Динамика показателей температуры в слуховом проходе на различных этапах исследования: а — группа I; б — группа II; в — группа III; Δ — p<0,01 уровень статистической значимости при сравнении между этапами исследования (критерий Вилкосона); * — p<0,05 — между группами (критерий Манна—Уитни)
Показатели испытуемых трех исследуемых групп различались по параметрам ВСР, большие значения SDNN, pNN50%, RMSSD, ТР регистрировались у испытуемых группы II (табл. 2). В данной группе SI меньше 50 у.е. отмечалось у 75% обследуемых. Наименьшие значения SDNN, pNN50%, RMSSD, ТР наблюдались у испытуемых группы III, где SI больше 150 у.е. регистрировалось у 62,5% обследованных. В группе I значения SI находились в пределах 50—150 у.е. в 75% случаев.
При воздействии холода у испытуемых всех групп отмечалось повышение значений SDNN и ТР. Анализ спектральной мощности ВСР выявил повышение HF, мс2, LF, мс2 у испытуемых группы I после пребывания на холоде. В группе II отмечалось повышение HF, мс2; у респондентов группы III повышались HF, мс2, LF, мс2, VLF, мс2. Через 5 минут после холода показатели ВСР у испытуемых групп I и II достигали значений, зарегистрированных до воздействия холода, а в группе III не восстанавливались до фоновых значений. Результаты анализа показателей ЧСС в каждой из трех групп в ситуации фоновой оценки не обнаружили различий в значениях. В группах I и III отмечалось снижение показателей ЧСС через 5 минут после пребывания на холоде. Различий в значениях показателей ЧСС у испытуемых группы II на различных этапах исследования обнаружено не было.
Анализ различий фоновых значений биоэлектрической активности головного мозга у испытуемых исследуемых групп показал, что у респондентов группы III обнаруживается статистически значимая более высокая СМ α-активность ЭЭГ в затылочных (О2, р=0,016, О1, р=0,013), центральных (С4, р=0,011, С3, р=0,011), лобных (F4, р=0,003, F3, р=0,001) и передневисочных (F8, р=0,001, F7, р=0,001) отделах обоих полушарий, в теменном отделе (P3, р=0,007) слева по сравнению с испытуемыми группы I, и теменных (Р4, р=0,015, Р3, р=0,011), лобных (F4, р=0,011, F3, р=0,015) отделах обоих полушарий, центральном (С3, р=0,015), заднетеменном (Т6, р=0,015) и передневисочном (F8, р=0,012) справа, по сравнению с испытуемыми группы II. Также более высокая СМ θ-активность регистрировалась в затылочных (O2, р=0,001, O1, р=0,005), теменном (P4, р=0,013), центральном (C4, р=0,001), лобном (F4, р=0,005), задневисочном (T6, р=0,013) и средневисочном (T4, р=0,011) отделах справа по сравнению с испытуемыми группы I, в затылочном (O2, р=0,003, O1, р=0,015), центральном (C3, р=0,012) слева, лобных (F4, р=0,012, F3, р=0,012) и височных справа (T6, р=0,015, T4, р=0,015, F8, р=0,015) по сравнению с испытуемыми группы II. Также у испытуемых группы II регистрировалась более низкая СМ β1-активность ЭЭГ в теменных отделах мозга по сравнению с испытуемыми двух других групп.
Таблица 2
Параметры вариабельности ритма сердца на различных этапах исследований, Ме (25; 75)
|
Показатель |
Группа I (N=12) |
Группа II (N=8) |
группа III (N=8) |
|||||||||
|
ФОН |
Холод |
Тепло через |
р, по |
ФОН |
Холод |
Тепло через |
р, по |
ФОН |
Холод |
Тепло через |
р, по |
|
|
1 |
2 |
3 |
1 |
2 |
3 |
1 |
2 |
3 |
||||
|
ЧСС |
72,6 (67,3;80,4) |
70,2 (65,2;76,5) |
67,1##1-3 (60,9;70,9) |
0,001 |
66,5 (59,1;73,8) |
70,4 (64,9;74,2) |
63,3##1-3 (59,3;65,3) |
0,091 |
81,1 (68,4;90,8) |
76,6 (71,3;79,3) |
70,7#1-3 (64,8;80,5) |
0,072 |
|
r MSSD, мс |
38,7 (34,9;58,3) |
63,4##1-2 (39,0;83,2) |
53,0 (42,0;78,7) |
0,016 |
68,1**I-II (46,8;79,6) |
83,4 (66,3;111,8) |
69,3 (58,1;89,5) |
0,072 |
25,2**I,II-III (18,1;35,5) |
53,4##1-2(45,9;71,5) |
46,7##1-3 (31,8;68,4) |
0,001 |
|
рNN50% |
19,1 (11,3;37,9) |
29,8 (15,2;53,7) |
29,9 (18,3;44,1) |
0,205 |
48,1**I-II (27,4;56,9) |
57,5 (44,5;62,1) |
46,4 (37,7;55,6) |
0,416 |
4,2**I,II-III (1,1;16,8) |
31,4##1-2 (20,2;43,9) |
18,9##1-3 (5,9;41,7) |
0,001 |
|
SDNN, мс |
55,7 (43,8;72,9) |
83,1##1-2 (65,9;103,5) |
71,6 (53,9;93,6) |
0,004 |
82,5 (63,7;100,8) |
95,9##1-2 (85,8;139,6) |
84,5 (64,4;115,9) |
0,043 |
38,7**I,II-III (32,7;44,7) |
76,4##1-2 (59,5;100,5) |
63,6##1-3 (42,0;106,8) |
0,002 |
|
SI, у.е. |
90,2 (48,7;122,4) |
36,9 (22,5;64,0) |
49,2##1-3 (30,6;66,9) |
0,008 |
37,7 (26,7;59,6) |
26,7 (11,6;37,8) |
32,8##1-3(15,6;46,3) |
0,007 |
168,2**I,II-III (116,2;277,8) |
51,2##1-2 (28,9;77,8) |
60,2##1-3 (23,4;130,1) |
0,002 |
|
ТР, мс2 х1000 |
2,9 (2,3;3,6) |
5,4##1-2 (2,7;11,8) |
4,8 (2,9;8,7) |
0,038 |
5,9 (3,4;6,7) |
7,2##1-2 (5,3;14,5) |
5,6 (3,7;12,6) |
0,011 |
1,3**I,II-III (1,2;1,8) |
5,3##1-2 (3,2;9,0) |
4,7##1-3 (2,4;7,5) |
0,002 |
|
HF, мс2 х1000 |
0,6 (0,5;1,1) |
1,3##1-2 (0,7;2,2) |
0,8 (0,6;1,8) |
0,008 |
1,6 (0,8;1,8) |
2,9##1-2 (2,2;3,7) |
1,5 (1,1;1,9) |
0,009 |
0,4**I,II-III (0,2;0,5) |
1,2##1-2(0,8;3,0) |
0,7##1-3 (0,5;1,5) |
0,004 |
|
LF, мс2 х1000 |
1,0 (0,7;1,3) |
2,7#1-2 (1,1;4,4) |
1,5 (0,9;3,6) |
0,027 |
1,7**I-II (1,2;2,1) |
2,2 (1,4;4,0) |
1,8 (1,1;2,8) |
0,194 |
0,5**I-III (0,3;0,8) |
1,5##1-2 (1,1;3,2) |
1,2##1-3(0,9;2,5) |
0,002 |
|
VLF, мс2 х1000 |
0,8 (0,5;1,1) |
0,7 (0,4;1,9) |
1,0 (0,7;1,3) |
0,778 |
1,1 (0,8;1,6) |
1,1 (0,6;1,6) |
1,0 (0,7;1,9) |
0,417 |
0,3**I,II-II (0,2;0,3) |
0,7##1-2 (0,5;1,1) |
0,5##1-3 (0,2;1,3) |
0,030 |
Примечание: «#» — р <0,05; «##» — p <0,01 уровень статистической значимости при попарном сравнении между этапами исследования (критерий Вилкосона); «**» — р<0,01 уровень статистической значимости между группами (критерии Манна—Уитни).

Рис. 4. Изменение спектральная мощности активности (4—13 Гц) ЭЭГ у испытуемых при воздействии холода: А — группа I, Б — группа II; Δ — статистически значимое повышение спектральной мощности ЭЭГ в диапазоне 4—13 Гц
У испытуемых группы II в ответ на воздействие холода статистически значимо повышалась СМ θ-активность в височных отделах обоих полушарий (F7, p=0,016, F8, p=0,016, T3, p=0,016, T4, p=0,016, T5, p=0,016, T6, p=0,016) (рис. 4) и лобном отделе (F4, p=0,017). У испытуемых группы III в ответ на воздействие холода не было выявлено статистически значимых изменений СМ α- и θ-активности. Показатели СМ β1-активности были оставлены за пределами анализа, поскольку динамика данных показателей обнаруживает взаимосвязь с напряжением мышц головы.
Обсуждение результатов
Теория нейровисцеральной интеграции (Thayer, Lane) предполагает, что центральная и вегетативная нервная системы взаимосвязаны между собой, и информация передается в двух направлениях [Smith, 2017]. Имеются доказательства того, что префронтальная кора участвует в модуляции работы сердца через блуждающий нерв, а ВСР является индикатором работы вегетативной нервной системы, связанной с когнитивной гибкостью при выполнении задач на внимание и рабочую память [Alba, 2019; Hansen, 2009].
В группе I были выявлены достаточно высокие значения ТР, RMSSD, pNN50%, а значения SI — в пределах от 50 до 150 у.е. при фоновой оценке, что свидетельствует о сбалансированном влияние симпатической и парасимпатической нервной системы на регуляцию ритма сердца у испытуемых данной группы. Сравнительный анализ показателей ТР, RMSSD, pNN50% и SI свидетельствует о наибольших значениях ТР, RMSSD, pNN50% и наиболее низких значениях SI (ниже 50 у.е.) у испытуемых группы II по сравнению с группами I и III. В данном случае можно говорить о преобладании в регуляции ритма сердца активности парасимпатической нервной системы. По мнению ряда авторов, повышенная вагусная активность может быть связана со слабой централизацией управления сердечным ритмом и преобладанием автономного контура регуляции [Шлык, 2013]. Респонденты группы III отличались по параметрам ВСР более низкими значениями ТР, RMSSD, pNN50%, HF, мс2, LF, мс2, VLF, мс2 и высокими значения SI, что свидетельствует о преобладании в регуляции ритма сердца активности симпатической нервной системы и участии центральных регулирующих систем. Более низкая амплитуда вазомоторных (LF) и низкочастотных (VLF) волн в спектре ВСР указывает на существенное напряжение стволового сосудодвигательного центра и надсегментарных влияний [Шлык, 2013].
Согласно поливагальной теории Porges S.W., существует связь между тонусом блуждающего нерва и функционированием систем саморегуляции внимания и эмоций [Thayer, 2009]. Он предположил, что более высокие уровни тонуса блуждающего нерва в покое связаны с хорошими показателями внимания. С данной теорией согласуются полученные у испытуемых группы I и III результаты, свидетельствующие о более высоких значениях ВСР при средней скорости выполнения теста в группе I по сравнению с группой III, у испытуемых которой отмечалась низкая скорость выполнения теста. Низкая ВСР предсказывает сверхбдительность и неэффективное распределение внимания и когнитивных ресурсов [Thayer, 2005].
Однако в группе II, у испытуемых которой регистрировались самые высокие показатели ВСР, наблюдалось большее количество ошибок при выполнении теста. В нескольких исследованиях была обнаружена положительная корреляция между более высокими уровнями тонуса блуждающего нерва и количеством ошибок при выполнении когнитивной задачи, а также между показателями ВСР и функциями внимания, как в период покоя, так и во время выполнения задач [Duschek, 2009]. Duschek и соавт. предположили, что сложные задачи или задачи, выполняемые в сжатые сроки, требуют более высоких энергетических ресурсов и таким образом производительность может в значительной степени зависеть от активации метаболических ресурсов [Duschek, 2009]. Повышение тонуса блуждающего нерва связано с трофотропной функцией и поэтому не оказывает сколь-нибудь существенного влияния на производительность работы в данном случае. Можно считать, что снижение тонуса блуждающего нерва во время выполнения задания помогает установить эрготропное физиологическое состояние, необходимое для удовлетворения метаболических требований, и способствует оптимизации когнитивного функционирования [Duschek, 2009]. В исследовании с использованием фармакологических средств, которые позволяют усилить периферическое и центральное симпатическое влияние, было показано улучшение памяти; напротив, включение в работу антагонистов β-адренорецепторов, которые ослабляют симпатическую активность, может приводить к ухудшению памяти [Critchley, 2013]. Вероятно, в группе II большее количество ошибок может быть связано с недостаточной активностью симпатической нервной системы.
Результаты анализа показателей испытуемых в фоновом исследовании указывают на различия в биоэлектрической активности головного мозга — у испытуемых группы III по сравнению с испытуемыми группы I и II регистрировались статистически большие значения СМ α- и θ-активности ЭЭГ, что может отражать снижение ингибирующего влияния со стороны коры на подкорковые структуры, и, следовательно, в механизмах саморегуляции мозга преобладают гипоталамо-диэнцефальные влияния [Сороко, 2009]. Thayer и Lane предположили, что преобладание симпатической нервной системы при гипоактивации префронтальной коры будет способствовать растормаживанию миндалевидного тела, что приводит к снижению ВСР [Thayer, 2009] и, возможно, к снижению когнитивной гибкости.
В зависимости от исходных значений выявлены особенности направленности изменения активности регуляторных систем при воздействии холода. При воздействии холода организм реагирует снижением температуры поверхности тела. Холодовой стресс вызывает периферическое сужение сосудов, опосредованное активностью симпатической нервной системы.Уменьшение периферического кровотока снижает конвективный теплообмен между ядром тела и оболочкой (кожей, подкожно-жировой клетчаткой и скелетными мышцами) и увеличивает изоляцию [Nutritional needs in, 1996]. Таким образом, во время воздействия холода защита ядра тела происходит за счет снижения температуры кожи.
У испытуемых группы III снижение температуры происходило быстрее; так, через 5 минут нахождения в холодовой камере Тсл была статистически ниже по сравнению с фоновым обследованием (тепло) и с Тсл испытуемых группы I, что, вероятно, возникало из-за большого периферического сужения сосудов поверхности тела. Через 10 минут нахождения в холодовой камере у испытуемых всех обследованных групп регистрировались более низкие Тк и Тсл по сравнению с фоновым обследованием; при этом в группе III отмечалась более низкая Тк по сравнению с группой I. Акклиматизация к холоду приводит к уменьшению сужения сосудов, повышению температуры кожи, задержке включения механизма дрожи, замедлению высвобождения гормонов стресса [Mäkinen, 2006] с целью предохранения поверхностных тканей от повреждения холодом и обеспечения работоспособности.
Приспособительные реакции организма на холод обеспечиваются работой различных теплорегуляционных механизмов. В группе I при охлаждении повышалась парасимпатическая активность (RMSSD, pNN50, HF, мс2) и активность вазомоторного центра (LF, мс2). В группе II в ответ на холод наблюдалось повышение уровня HF, мс2, что может свидетельствовать о возрастании активности парасимпатической нервной системы. После акклиматизации к холоду наблюдается, как правило, снижение симпатической реакции и сопутствующее усиление активации блуждающего нерва [Mäkinen, 2008]. У испытуемых группы III при воздействии холода возрастала активность парасимпатической (RMSSD, HF, мc2) нервной системы, вазомоторного центра (LF, мc2) и центральных отделов вегетативной нервной системы (VLF, мс2). Для поддержания нормального уровня функционирования сердечно-сосудистой системы организм с центральным типом регуляции затрачивает больше усилий, нежели организм с автономным типом регуляции [Шлык, 2013]. Установлено, что у индивидов с центральным типом регуляции ритма сердца происходит напряжение адаптационных механизмов, приводящее к снижению резервных возможностей организма [Шлык, 2013]. После прекращения воздействия холода у испытуемых групп I и II параметры ВСР восстанавливались до исходных показателей в отличие от испытуемых группы III, где параметры ВСР соответствовали значениям при воздействии холода.
Результаты анализа особенностей реагирования на холод указывают на различную динамику изменений биоэлектрической активности головного мозга. В группе I повышалась СМ θ-активность в височной области справа, центральной, в лобной, а также в передневисочном отделе слева. Также повышалась СМ α-активность в передневисочных и лобных отделах. При воздействии холода происходит активация гипоталамо-диэнцефальных структур головного мозга. В исследованиях Fechir и соавт. отмечено, что измерения ПЭТ-ФДГ показали связанную с охлаждением активацию в мозжечке и стволе мозга, а также деактивацию островка справа и передней поясной коры [Fechir, 2010]. Результаты анализа показателей испытуемых группы I позволяют предположить, что снятие тормозного влияния префронтальной, передней поясной коры и островка преимущественно справа на нижележащие структуры головного мозга при воздействии холода будет усиливать активность парасимпатической нервной системы и вазомоторного центра. Такой вывод согласуется с данными других исследований, указывающих на специализацию полушарий головного мозга в регуляции вегетативной нервной системы, где правое полушарие преимущественно связано с активностью симпатической нервной системы, тогда как левое полушарие — с парасимпатической [Guo, 2016]. У испытуемых группы II в ответ на холод повышалась СМ θ-активность в височных отделах обоих полушарий и лобном отделе справа. Вероятно, различия в реакции на воздействие холода между испытуемыми группы II и группы I определяются уровнем активации гипоталамо-диэнцефальных структур мозга. В группе III на фоне высокой активности гипоталамо-диэнцефальных структур головного мозга в ответ на холод изменение биоэлектрической активности не отмечалось.
Заключение
Таким образом, выявлено три варианта адаптационно-приспособительных реакций у молодых мужчин в ответ на воздействие низких температур (-200С), проявляющихся, в том числе, в точности работы произвольного внимания. У испытуемых группы I, отличающихся высокой точностью выполнения задания и средней скоростью обработки информации, отмечался более оптимальный режим функционирования систем организма в ответ на холод, что проявлялось в умеренном снижении температуры поверхности кожи, активацией парасимпатической нервной системы и вазомоторного центра, активацией гипоталамо-диэнцефальных структур головного мозга и снижением тормозного влияния префронтальной коры преимущественно справа (θ- и α-диапазоны ЭЭГ в лобных и передневисочных отделах обоих полушарий и височных отделах справа). У испытуемых группы II, показавших среднюю скорость выполнения с большим по сравнению с испытуемыми других групп количеством ошибок, наблюдалась приспособительная реакция на холод с недостаточной мобилизацией ресурсов, что отражалось в меньшем снижении температуры поверхности кожи, повышением активности парасимпатической нервной системы и увеличением активности подкорковых структур мозга (θ-активности ЭЭГ в височных отделах обоих полушарий и в лобном отделе справа). И наконец, у испытуемых группы III, характеризовавшихся низкой скоростью выполнения заданий теста, но достаточно высокой точностью, отмечалось напряжение адаптационно-приспособительных механизмов в ответ на холод, что проявлялось в выраженном снижении температуры поверхности кожи, усиленном ответе со стороны вегетативной нервной системы в виде повышения как парасимпатических, так и симпатических влияний (HF, LF, VLF) на регуляцию ритма сердца и сохранении высокой активности гипоталамо-диэнцефальных структур головного мозга. Таким образом, в условиях Севера выявлены индивидуальные различия в приспособительных реакциях в ответ на холод, связанные с разными нейрофизиологическими механизмами мобилизации функциональных систем и отражающиеся на эффективности работы произвольного внимания.
Литература
- Иванников В.А., Шляпников В.Н. Особенности волевой регуляции у представителей разных этнокультурных групп // Экспериментальная психология. 2019. Том 12. № 1. С. 70—84.
- Кривоногова Е.В. Когнитивный вызванный потенциал р300 у школьников 16—17 лет, проживающих в регионах арктической зоны РФ // Журнал медико-биологических исследований. 2020. Том 8. № 4. С. 360—367.
- Максимов А.Л., Аверьянова И.В. Особенности гемодинамики и вариабельности сердечного ритма у юношей-европеоидов при проведении активной ортостатической пробы // Экология человека. 2021. № 1. С. 22—31.
- Марков А.Л., Солонин Ю.Г., Бойко Е.Р. Влияние метеорологических параметров на вегетативную регуляцию ритма сердца у жителей европейского севера: индивидуальный контроль // Вестник Тверского государственного университета. Серия: Биология и экология. 2018. № 1. С. 21—29.
- Медведев В.И. Взаимодействие физиологических и психических механизмов в процессе адаптации // Физиология человека. 1998. Том 24. № 4. С. 7—13.
- Солонин Ю.Г. Исследования по широтной физиологии (обзор) // Журнал медико-биологических исследований. 2019. Том 7. № 2. С. 228—239. DOI: 10.17238/issn2542-1298.2019.7.2.228
- Сороко С.И., Андреева С.С., Бекшаев С.С. Перестройки параметров электроэнцефалограммы у детей — жителей о. Новая Земля // Вестник Северо-Восточного научного центра ДВО РАН. 2009. № 2. С. 49—59
- Шлык Н.И., Зуфарова Э.И. Нормативы показателей вариабельности сердечного ритма у исследуемых 16—21 года с разными преобладающими типами вегетативной регуляции // Вестник Удмуртского университета. Серия Биология. Науки о Земле. 2013. № 4. С. 96—105.
- Ясюкова Л.А. Оптимизация обучения и развития детей с ММД. Диагностика и компенсация минимальных мозговых дисфункций: метод. Руководство. СПб.: ГП «ИМАТОН», 1997. 80 с.
- Alba G., Vila J., Rey B., Montoya P., Muñoz M.Á. The relationship between heart rate variability and electroencephalography functional connectivity variability is associated with cognitive flexibility // Front. Hum. Neurosci. 2019. Vol. 13. P. 64. DOI: 10.3389/fnhum.2019.00064
- Critchley H., Eccles J., Garfinkel S. Interaction between cognition, emotion, and the autonomic nervous system // Handbook of clinical neurology. 2013. Vol. 117. P. 59—77. DOI: 10.1016/B978-0-444-53491- 0.00006-7
- Duschek S., Muckenthaler M., Werner N., Reyes del Paso G.A. Relationships between features of autonomic cardiovascular control and cognitive performance // Biol. Psychol. 2009. Vol. 81. P.110—117. DOI: 10.1016/j.biopsycho.2009.03.003
- Fechir M., Klega A., Buchholz H.G., Pfeifer N., Balon S., Schlereth T., Geber C., Breimhorst M., Maihofner C., Birklein F., Schreckenberger M. Cortical control of thermoregulatory sympathetic activation // Eur J. Neurosci. 2010. Vol.31. P. 2101—2111. DOI: 10.1111/j.1460-9568.2010.07243.x
- Guo C.C., Sturm V.E., Zhou J., Gennatas E.D., Trujillo A.J., Hua A.Y., Crawford R., Stables L., Kramer J.H., Rankin K., Levenson R.W., Rosen H.J., Miller B.L., Seeley W.W. Dominant hemisphere lateralization of cortical parasympathetic control as revealed by frontotemporal dementia // Proc. Natl. Acad. Sci USA. 2016. Vol. 113. № 17: E2430-9. DOI: 10.1073/pnas.1509184113
- Hansen A.L., Johnsen B.H., Thayer J.F. Relationship between heart rate variability and cognitive function during threat of shock // Anxiety Stress Coping. 2009. Vol. 22. P. 77—89. DOI: 10.1080/10615800802272251
- Mäkinen T.M., Palinkas L A., Reeves D.L., Pääkkönen T., Rintamäki H., Leppäluoto J., Hassi J. Effect of repeated exposures to cold on cognitive performance in humans // Physiol. Behav. 2006. Vol. 87. P. 166— 176. DOI: 10.1016/j.physbeh.2005.09.015
- Mäkinen T.M., Mäntysaari M., Pääkkönen T., Jokelainen J., Palinkas L.A., Hassi J., Leppäluoto J., Tahvanainen K., Rintamäki H. Autonomic nervous function during whole-body cold exposure before and after cold acclimation // Aviat. Space Environ Med. 2008. Vol. 79. № 9. P. 875—882. DOI: 10.3357/ asem.2235.2008
- Nutritional needs in cold and in high-altitude environments: applications for military personnel in field operations. Marriott B.M., Carlson S.J. editors. Washington (DC): National academies press (US), 1996. PMID: 25121290.
- Posner M., Rothbart M., Ghassemzadeh H. Developing attention in typical children related to disabilities // Handbook of clinical neurology. 2020. Vol. 173. P. 215—223. DOI: 10.1016/B978-0-444-64150-2.00019-8
- Smith R., Thayer J.F., Khalsa S.S., Lane R.D. The hierarchical basis of neurovisceral integration // Neurosci. Biobehav. Rev. 2017. Vol. 75. P. 274—296. DOI: 10.1016/j.neubiorev.2017.02.003
- Spangler D.P., Cox K.R., Thayer J.F., Brooks J.R., Friedman B.H. Interplay between state anxiety, heart rate variability, and cognition: An ex-Gaussian analysis of response times // Int J Psychophysiol. 2021. Vol. 159. P. 60—70. DOI: 10.1016/j.ijpsycho.2020.08.018
- Thayer J.F., Brosschot J.F. Psychosomatics and psychopathology: looking up and down from the brain // Psychoneuroendocrinology. 2005. Vol. 30. № 10. P. 1050—1058. DOI: 10.1016/j.psyneuen.2005.04.014
- Thayer J.F., Lane R.D. Claude Bernard and the heart—brain connection: further elaboration of a model of neurovisceral integration // Neurosci. Biobehav. Rev. 2009. Vol. 33. P. 81—88. DOI: 10.1016/j. neubiorev.2008.08.004
Информация об авторах
Метрики
Просмотров
Всего: 827
В прошлом месяце: 30
В текущем месяце: 0
Скачиваний
Всего: 204
В прошлом месяце: 7
В текущем месяце: 0