Экспериментальная психология
2018. Том 11. № 3. С. 51–62
doi:10.17759/exppsy.2018110304
ISSN: 2072-7593 / 2311-7036 (online)
Роль категориальной идентичности стимулов в возникновении эффекта «пропусков при продолжении поиска»
Аннотация
Общая информация
Ключевые слова: зрительный поиск, зрительное внимание, перцептивная установка, эффект «пропусков при продолжении поиска»
Рубрика издания: Когнитивная психология
Тип материала: научная статья
DOI: https://doi.org/10.17759/exppsy.2018110304
Для цитаты: Ланина А.А., Горбунова Е.С. Роль категориальной идентичности стимулов в возникновении эффекта «пропусков при продолжении поиска» // Экспериментальная психология. 2018. Том 11. № 3. С. 51–62. DOI: 10.17759/exppsy.2018110304
Полный текст
Введение
Эффект «пропусков при продолжении поиска» (ПППП) представляет тобой снижение успешности нахождения второго целевого стимула после успешного нахождения первого целевого стимула в задаче зрительного поиска (Adamo, Cain, Mitroff, 2013). Изначально данный феномен был обнаружен врачами-рентгенологами и получил название «насыщение поиска» (см. например: Tuddenham, 1962), поскольку его первое теоретическое объяснение заключалось в том, что врач после нахождения на рентгеновском снимке какой-либо аномалии довольствуется результатом и прекращает поиск других возможных нарушений. Тем не менее, более поздние исследования показывают, например, что время сканирования дисплея не зависит от наличия одного или нескольких стимулов, что не соответствует предполагаемому паттерну результатов для гипотезы «насыщения» (Berbaum, Franken, Dorfman, Rooholamini, Coffman, Cornell, Cragg, Galvin, Honda, Kao, et al, 1991). Таким образом, «насыщение» если и является причиной ПППП, то явно не самой главной. В связи с этим были предложены две альтернативные теории, которые на данный момент являются основными.
дложены две альтернативные теории, которые на данный момент являются основными. Первая из этих теорий связывает пропуск второго целевого стимула с истощением ресурсов внимания и/или рабочей памяти. Согласно теории истощения ресурсов, на обработку первого целевого стимула тратится существенное количество ресурсов внимания и/или рабочей памяти, в связи с чем на поиск второго целевого стимула ресурсов уже не остается. Данная теория нашла подкрепление в ряде исследований. Например, амплитуда эффекта ПППП возрастает при увеличении числа расположенных рядом с целевым стимулом дистракторов, которые вызывают дополнительную нагрузку на ограниченные ресурсы внимания (Adamo, Cain, Mitroff, 2015). Напротив, вероятность нахождения второго целевого стимула увеличивается, если первый найденный стимул пропадает с экрана или становится более заметным относительно фона (Cain, Mitroff, 2013). Вероятность пропуска второго целевого стимула также снижается, когда после обнаружения первого целевого стимула предъявляется несколько пустых проб, а уже потом реализуется дальнейший поиск (Cain, Biggs, Darling, Mitroff, 2014). Наконец, эффект «пропусков при продолжении поиска» обнаруживает поразительное сходство с другим известным перцептивным феноменом — эффектом «мигания внимания», возникновение которого также зачастую связывается с истощением ресурсов рабочей памяти (Adamo, Cain, Mitroff, 2013).
Тем не менее, результаты наших исследований с одновременным выполнением задач на пространственную рабочую память и зрительный поиск двух целевых стимулов не выявили значимой интерференции между задачами (Горбунова, 2017). Имеются основания полагать, что если истощение ресурсов рабочей памяти каким-то образом причастно к пропуску второго целевого стимула, то речь идет скорее об объектной подсистеме, осуществляющей хранение объектных репрезентаций стимулов.
Альтернативное объяснение феномена ПППП предлагается в рамках теории перцептивной установки. Согласно этой теории, после нахождения первого целевого стимула создается своего рода «перцептивное смещение» — испытуемый склонен будет различать стимулы, сходные с первым найденным, и пропускать стимулы, отличающиеся от него. Например, после нахождения на рентгене одного перелома врач с высокой вероятностью обнаружит другой, а опухоль с высокой вероятностью пропустит (Berbaum, Franken, Dorfman, Rooholamini, Coffman, Cornell, Cragg, Galvin, Honda, Kao, et al, 1991). Возможные механизмы этого перцептивного смещения могут подразумевать как восходящий прайминг со стороны стимулов в зрительном поле, так и нисходящее направление внимания (guidance) со стороны репрезентаций в рабочей памяти. В обоих случаях нахождение первого целевого стимула, вероятно, ведет к понижению порогов обнаружения для других перцептивно сходных стимулов (Kristjánsson, Campana, 2017). Исследователи, работающие в рамках данного когнитивного направления, обнаруживают большое количество эмпирических данных в пользу теории перцептивной установки. В частности, эффект ПППП уменьшается при увеличении количества общих перцептивных признаков (таких как форма, размер, цвет, ориентация) у двух целевых стимулов (Gorbunova, 2017). Помимо этого, результаты исследований стандартного зрительного поиска с одним целевым стимулом показывают, что дистракторы имеют большее влияние на эффективность зрительного поиска, когда они являются перцептивно сходными с целевым стимулом (Duncan, Humphreys, 1989).
Тем не менее, результаты первых исследований ПППП в лабораторных условиях свидетельствуют о том, что эффект наблюдается даже для полностью идентичных стимулов — двух букв «Т» одинакового контраста (Fleck, Samei, Mitroff, 2010). Однако поскольку в этом исследовании варьировалась ориентация стимулов, вопрос об их полной идентичности остается открытым, поскольку ориентация стимула в пространстве является базовым перцептивным признаком. Более существенным аргументом являются результаты исследования Биггса и коллег с использованием big data, проведенного на базе игры «Сканер в аэропорту», в которой необходимо найти на изображениях как будто бы провозимого пассажирами багажа запрещенные предметы (Biggs, Adamo, Dowd, Mitroff, 2015). В условии с двумя целевыми стимулами они могли иметь перцептивное сходство (одинаковый цвет, например, два синих объекта) либо категориальное (одинаковая функция или категория, например, пистолет и пули). Было обнаружено, что категориальное сходство вносит существенный вклад в возникновение ПППП, в то время как роль перцептивного сходства не столь велика при условии контроля категориального. Тем не менее, стоит отметить, что эти результаты могут быть связаны с ограничением перцептивного анализа одним признаком — цветом, а также, в ряде проб, вероятно, возможным совместным влиянием перцептивного и категориального сходства.
Также следует отметить, что две основные теории ПППП — истощение ресурсов и перцептивная установка — необязательно являются конкурирующими. Имеются основания предполагать, что репрезентация первого целевого стимула одновременно истощает ресурсы рабочей памяти в связи с необходимостью ее хранения и направляет внимание на поиск сходных стимулов. Помимо этого, перцептивная установка может выступать в качестве своеобразной адаптации к ситуации ограниченности ресурсов памяти.
В рамках данного исследования проводится экспериментальная проверка предположения о роли категориального сходства стимулов в возникновении эффекта ПППП, но только в отношении идентичности стимулов. Мы предполагаем, что данный эффект будет снижаться при предъявлении идентичных стимулов. Изучение причин возникновения эффекта ПППП имеет немаловажное значение не только для дальнейшей разработки моделей ПППП, но и для расширения теоретических представлений о процессе решения задачи зрительного поиска в целом, а также о роли категорий и механизмах направления внимания при решении данной задачи — в частности.
Цель нашего исследования состоит в изучении роли категориальной идентичности стимулов в возникновении эффекта ПППП.
Основная гипотеза исследования: эффект ПППП зависит от категориальной идентичности целевых стимулов, что выражается в большем количестве верных ответов и меньшем времени реакции в отношении второго целевого стимула после успешного нахождения первого целевого стимула в случае идентичных целевых стимулов по сравнению с условием неидентичных стимулов.
Методика
Испытуемые
В эксперименте приняли участие 22 человека в возрасте от 17 до 20 лет (M = 18,73), один мужского пола и 21 женского, студенты департамента психологии НИУ ВШЭ. Все участники эксперимента имели нормальное или скорректированное до нормального зрение и являлись наивными испытуемыми по отношению к гипотезам эксперимента.
Аппаратура
Для предъявления стимулов был использован компьютер Pentium Dual Core CPU E6500 и монитор LACIE electron 19 blue III, разрешение экрана — 1024 x 768, частота обновления — 85 Гц. Расстояние от испытуемого до экрана составляло 40 см. В качестве программного обеспечения использовалась программа Psychopy v. 1.82.01, операционная система — Ubuntu. Ответы испытуемых регистрировались с помощью стандартной компьютерной мыши и клавиатуры.
Процедура
В качестве стимулов использовались арабские цифры: 1, 2, 4, 5, 6, 7, 8, 9. Цифра 3 не использовалась по причине лучшей по сравнению с другими цифрами различимости. Размер каждой цифры составлял 0,72 х 1,57 угловых градусов. Задача первой группы испытуемых (50% от всей выборки) состояла в поиске четных цифр среди нечетных, а другой группы — нечетных цифр среди четных. При этом целевых стимулов могло быть два (хорошо заметный и плохо различимый), один (хорошо или плохо различимый) или ни одного. Под «различимостью» в данном случае имеется в виду отличие стимула от фона по контрасту. Стимулы предъявлялись на сером фоне, rgb: [128, 128, 128]. Целевые стимулы и дистракторы имели разные оттенки и тем самым разную степень отличимости от фона. Стимулы, выступающие в качестве дистракторов, были следующих цветов (по rgb): [70, 70, 70]; [90, 90, 90] и [105, 105, 105]. Дистракторы разных цветов равномерно распределялись в каждой пробе. Стимулы, выступающие в качестве целевых, были следующих цветов (по rgb): [70, 70, 70] (хорошо различимый целевой стимул) и [105, 105, 105] (плохо различимый целевой стимул). В каждой пробе на экране предъявлялось 20 стимулов (данное количество объектов было выбрано по результатам предварительной апробации методики для получения эффекта «пропусков при продолжении поиска»). Стимулы были случайным образом распределены по экрану с помощью методики невидимой «решетки» размером 10*6 ячеек с произвольным размещением внутри клеток этой решетки. Также внизу экрана были расположены две кнопки — «ОК» и «НЕТ», которые предназначались для ответа испытуемого, угловой размер — 6,85° х 4,32°. Пример стимульного материала приведен на рис. 1.
Основная серия эксперимента состояла из 444 проб. В 144 пробах испытуемому предъявлялось два целевых стимула (один хорошо различимый и один плохо различимый), в 100 пробах присутствовал один плохо различимый целевой стимул, в 100 пробах присутствовал один хорошо различимый целевой стимул, в 100 пробах целевые стимулы отсутствовали. Из 144 проб с двумя целевыми стимулами в 72 пробах стимулы были одинаковыми (например, 2 и 2), а в 72 пробах — разными (например, 2 и 4); стимулы (т. е. конкретные цифры) и их сочетания (для разных цифр) были распределены равномерно.
Задача испытуемого состояла в том, чтобы обнаружить все целевые стимулы, либо дать отчет об их отсутствии. Ответ давался с помощью щелчка мышью. При нахождении двух целевых стимулов испытуемый должен был последовательно осуществить щелчок мышью на каждый из них. При нахождении одного целевого стимула испытуемый должен был нажать сначала на стимул, а далее на кнопку «ОК». В случае отсутствия целевых стимулов испытуемый должен был дважды осуществить клик мышью на кнопку «НЕТ». Последовательность предъявления проб была рандомизирована. Время каждой пробы составляло 20 с; если испытуемый не находил целевого стимула в течение этого времени, проба заканчивалась. Для перехода к каждой следующей пробе испытуемый нажимал на пробел. Для ознакомления с процедурой эксперимента перед основной серией испытуемый проходил тренировочную серию из 8 проб.
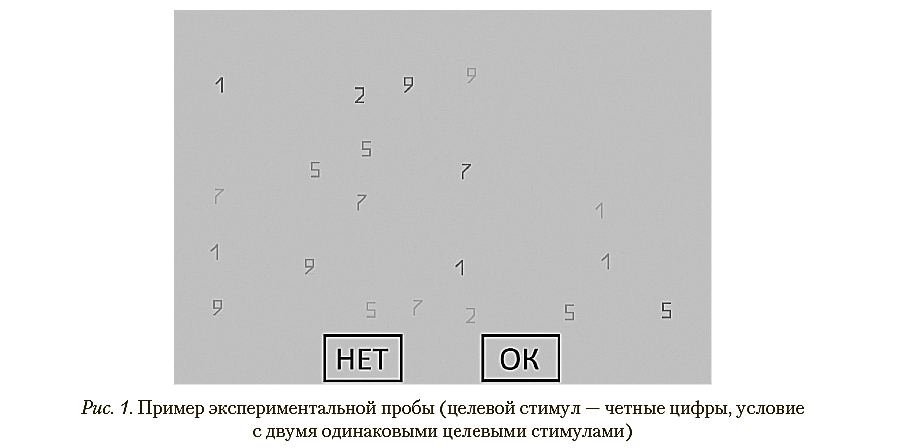
Обработка результатов
Сравнивалась успешность нахождения второго целевого стимула после успешного обнаружения первого целевого стимула, а также время первого и второго щелчка мышью в условиях с двумя одинаковыми целевыми стимулами и с двумя разными целевыми стимулами, а также в условии с одним плохо различимым целевым стимулом. Условия с отсутствием целевых стимулов и с одним хорошо различимым целевым стимулом в анализе не учитывались, поскольку не являются информативными в контексте выдвигаемых гипотез, и исходно вводились в качестве «проб-ловушек», однако данные по ним приводятся в таблицах ниже. В условии с двумя целевыми стимулами в качестве показателя точности ответов использовался процент верно обнаруженных плохо различимых целевых стимулов при условии верно обнаруженных хорошо различимых стимулов, при этом анализу подлежали только те пробы, в которых хорошо различимый стимул был верно обнаружен первым (это соответствует стандартной процедуре подсчета данных в экспериментах с ПППП (см. нгшример: Fleck, Samei, Mitroff, 2010). Для условия с одним плохо различимым целевым стимулом в качестве показателя точности использовался процент верно обнаруженных плохо различимых целевых стимулов при условии последующего нажатия на кнопку «НЕТ», соответствовавшего отчету о том, что найденный стимул является единственным. Данные обрабатывались с помощью SPSS 22.0. В качестве метода анализа данных был использован однофакторный дисперсионный анализ (ANOVA) с повторными измерениями. Были использованы парные сравнения (с поправкой на множественные сравнения Бонферрони—Холма) для сопоставления разных условий. Тест Маучли выявил значимое отклонение от сферичности для показателя процента верных ответов ( W = 0,581; p = 0,004), а также для времени второго клика мыши ( W = 0,656; p = 0,015), поэтому для этих измерений была использована поправка Гринхауса—Гейссера. Для времени первого клика мыши значимого отклонения от сферичности выявлено не было ( W" = 0,965; p = 0,698). Также произведена оценка размера эффекта методом ц2.
Результаты1
Процент верных ответов
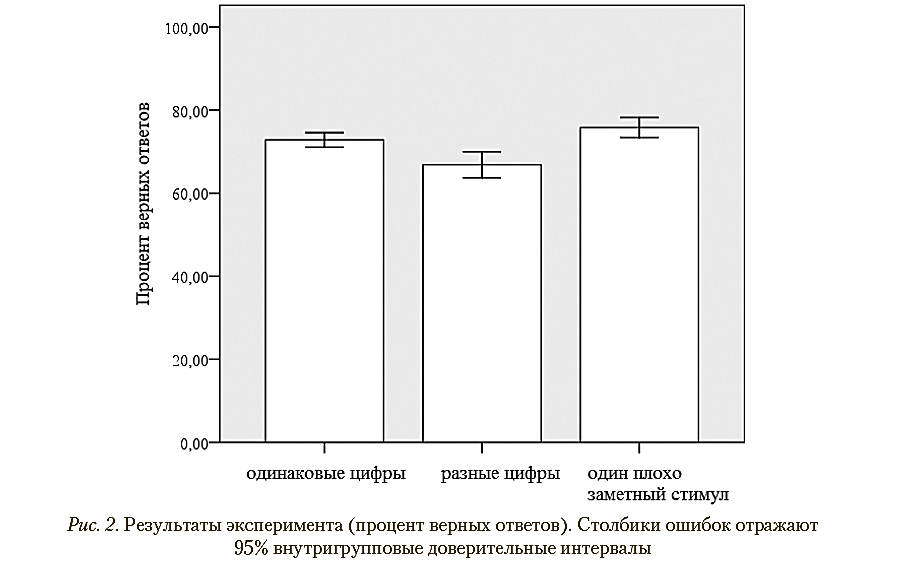
Дисперсионный анализ выявил значимое влияние фактора типа стимула: F (1, 30) = = 9,69; p = 0,002; nG = 0,316. Попарные сравнения (с поправками Бонферрони—Холма) выявили значимые различия между условиями с двумя одинаковыми целевыми стимулами и двумя разными целевыми стимулами (p = 0,023), между условиями с двумя одинаковыми стимулами и одним плохо различимым целевым стимулом (p = 0,034) и между условиями с двумя разными целевыми стимулами и одним целевым стимулом (p = 0,006). Результаты представлены в табл. 1, а также в графической форме на рис. 2.
Таблица 1
Результаты эксперимента (процент верных ответов)
|
|
Экспериментальные условия |
||||
|
Одинаковые цифры |
Разные цифры |
Один плохо различимый стимул |
Один хорошо различимый стимул |
Отсутствие целевых стимулов |
|
|
Среднее |
72,85 |
66,84 |
75,82 |
78,18 |
93,86 |
|
Стандартное отклонение |
17,99 |
19,71 |
16,49 |
16,56 |
5,9 |
Время первого щелчка мыши
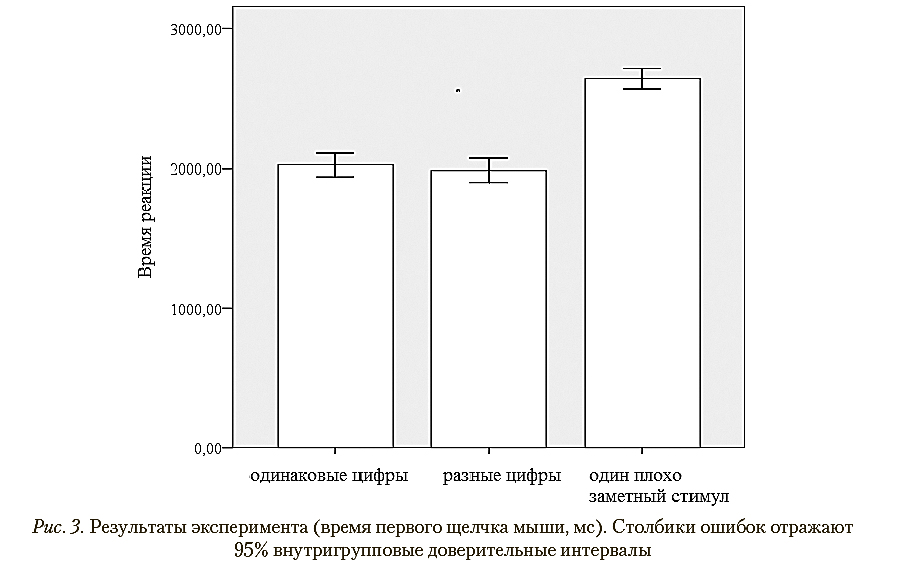
Дисперсионный анализ выявил значимое влияние фактора типа стимула: F (2, 42) = 59,76; p < 0,001; η2 = 0,740. Попарные сравнения (с поправками Бонферрони—
Холма) выявили значимые различия между условиями с двумя одинаковыми стимулами и одним плохо различимым целевым стимулом (р < 0,001) и между условиями с двумя разными целевыми стимулами и одним целевым стимулом (р < 0,001). Значимых различий между условиями с двумя одинаковыми целевыми стимулами и двумя разными целевыми стимулами обнаружено не было (р = 0,623). Результаты представлены в табл. 2, а также в графической форме на рис. 3.
Таблица 2
Результаты эксперимента (время первого щелчка мыши)
|
|
Экспериментальные условия |
||||
|
Одинаковые цифры |
Разные цифры |
Один плохо различимый стимул |
Один хорошо различимый стимул |
Отсутствие целевых стимулов |
|
|
Среднее |
2022,81 |
1986,05 |
2644,09 |
2597,61 |
4091,21 |
|
Стандартное отклонение |
259,2 |
462,62 |
467,43 |
450,05 |
908,25 |
Время второго щелчка мыши
Дисперсионный анализ выявил значимое влияние фактора типа стимула: F (1, 31) = 28,29; р < 0,001; nG = 0,574. Попарные сравнения (с поправками Бонферрони— Холма) выявили значимые различия между условиями с двумя одинаковыми стимулами и одним плохо различимым целевым стимулом (р < 0,001), между условиями с двумя разными целевыми стимулами и одним целевым стимулом (р = 0,002), а также между условиями с двумя одинаковыми целевыми стимулами и двумя разными целевыми стимулами (р < 0,001). Результаты представлены в табл. 3, а также в графической форме на рис. 4.
Таблица 3
Результаты эксперимента (время второго щелчка мыши)
|
|
Экспериментальные условия |
||||
|
Одинаковые цифры |
Разные цифры |
Один плохо различимый стимул |
Один хорошо различимый стимул |
Отсутствие целевых стимулов |
|
|
Среднее |
1353,28 |
1683,94 |
2070,81 |
2181,31 |
207,7 |
|
Стандартное отклонение |
320,24 |
294,51 |
648,12 |
698,14 |
47,66 |
Обсуждение результатов
Было обнаружено, что тип стимулов оказывает значимое влияние на вероятность обнаружения целевого объекта в задаче зрительного поиска множественных стимулов. В условии с двумя целевыми стимулами для двух разных цифр вероятность нахождения второй из них после успешного нахождения первой оказалась ниже по сравнению с условием с двумя одинаковыми цифрами. Тем не менее, эффект ПППП — снижение успешности нахождения второго целевого стимула после успешного нахождения первого — был обнаружен как в условии с одинаковыми, так и в условии с разными цифрами. Такого рода данные свидетельствуют, с одной стороны, в пользу влияния категориальной идентичности стимулов на возникновение эффекта ПППП, а с другой стороны, о возможном различии в механизмах работы перцептивной и категориальной установок, поскольку перцептивное сходство стимулов нивелирует эффект ПППП (Gorbunova, 2017). Помимо этого, следует отметить, что категориальное сходство в данном случае, с одной стороны, сводилось к идентичности стимулов, а с другой стороны, сочеталось с перцептивным сходством (одинаковые цифры
не только относятся к одной категории, но и имеют одинаковую форму). Хотя в рамках данного эксперимента была осуществлена попытка разделения категориального и перцептивного сходства (первый и второй целевой стимулы имели разный оттенок), очевидно, что перцептивное различие в данном случае было неполным. Вероятно, в качестве перспективы исследований можно назвать проведение дополнительных экспериментов с более полным разделением влияния этих двух факторов — перцептивного сходства и категориального. Еще одно возможное ограничение исследования может быть связано с гендерной неоднородностью выборки.
Время второго щелчка мыши (время нахождения второго целевого стимула) оказывается значимо меньшим для условия с двумя одинаковыми целевыми стимулами по сравнению с условием с двумя разными целевыми стимулами, что также подтверждает высказанные нами ранее предположения относительно роли категориальной идентичности двух целевых стимулов.
Время второго щелчка мыши оказывается наибольшим для условия с одним плохо различимым целевым стимулом. Этот результат аналогичен результатам, полученным нами ранее (Gorbunova, 2017), и, вероятно, связан с тем, что для условия с одним целевым стимулом второй щелчок мыши соответствует ответу о том, что второго целевого стимула не обнаружено. Ранее в классических исследованиях зрительного поиска было обнаружено, что время ответа для условия с отсутствием стимула оказывается большим по сравнению с условием с наличием стимула (Kwak, Dagenbach, Egeth, 1991).
Время первого щелчка мыши не отличается значимо для условий с двумя одинаковыми и двумя разными целевыми стимулами, однако оказывается значимо большим для условия с одним стимулом. Таким образом, время обнаружения хотя бы одного стимула в случае, когда их два, оказывается меньшим по сравнению со временем обнаружения единственного целевого стимула в зрительном поле. Поскольку местоположения стимулов в нашем эксперименте варьировались случайным образом, то можно предположить, что параметры времени обнаружения стимула взаимосвязаны с вероятностью нахождения единственного либо одного из двух целевых стимулов при сканировании зрительного поля. Существует также альтернативное объяснение полученных результатов. Поскольку мы использовали стандартную парадигму подсчета данных в экспериментах с ПППП (учитываются только те пробы, в которых первым был найден хорошо различимый стимул, и данные по ним сравниваются с условием одного плохо различимого целевого стимула), первый щелчок мыши для условия с двумя целевыми стимулами отражает процесс обнаружения хорошо различимого целевого стимула, а для условия с одним плохо различимым стимулом — обнаружения этого плохо различимого стимула соответственно. Таким образом, либо на обнаружение хорошо различимого целевого стимула тратится меньше ресурсов внимания по сравнению с плохо различимым целевым стимулом, либо внимание в первую очередь направляется на хорошо различимые стимулы (что, в свою очередь, также может быть связано с количеством ресурсов, затрачиваемых на обработку хорошо и плохо различимых стимулов).
Результаты исследования могут быть рассмотрены в контексте теории (перцептивной) установки, при условии ее расширения до теории перцептивной и категориальной установки. Первый найденный целевой стимул формирует установку на то, как должен выглядеть второй. При этом эта установка может быть как перцептивной (стимулы, сходные по зрительным признакам), так и категориальной (стимулы, принадлежащие к одной категории). При этом конкретный механизм категориальной установки, вероятно, связан с нисходящим направлением внимания: репрезентация первого целевого стимула в рабочей памяти способствует направлению внимания на стимулы той же категории. Существует также альтернативное объяснение, связывающее действие категориальной установки не с нисходящими, а с восходящими процессами. Ранее было обнаружено, что скорость обнаружения целевого стимула—цифры среди дистракторов — других цифр — зависит от числового расстояния между ними: чем оно больше, тем быстрее обнаруживается целевой стимул (Schwarz, Eiselt, 2012). Авторы работы связывают данный эффект со спецификой перцептивной деятельности на ранних этапах обработки информации.
Помимо этого, результаты данного эксперимента могут быть рассмотрены в контексте комбинированной (смешанной) теории, объединяющей в себе принципы теории установки и теории истощения ресурсов. Можно предположить, что либо репрезентация первого целевого стимула в рабочей памяти одновременно истощает ресурсы и создает установку на поиск последующих стимулов, либо сама установка является адаптацией к истощению ресурсов. В любом случае, первый целевой стимул способствует реконфигурации фильтра для поиска сходных объектов. Такое толкование согласуется с результатами некоторых экспериментов по изучению эффекта «мигания внимания» — феномена, обнаруживающего поразительное сходство с ПППП (см. например: Adamo, Cain, Mitroff, 2012). К примеру, было обнаружено, что в условиях быстрого последовательного предъявления зрительных стимулов увеличение сходства целевых стимулов и дистракторов приводит к повышению успешности опознания стимула-зонда, если он предъявляется сразу же после предъявления первого целевого стимула (Visser, Davis, Ohan, 2009).
Выводы
1. С целью проверки предположения о роли категориального сходства целевых стимулов в возникновении феномена ПППП был проведен эксперимент с предъявлением стимульного материала в виде цифр, в котором основной задачей испытуемых было нахождение целевых стимулов как хорошо, так и плохо различимых среди цифр-дистракторов. В условии с двумя целевыми стимулами цифры могли быть одинаковыми или разными. Было обнаружено, что данный эффект снижается при предъявлении сходных стимулов — одинаковых цифр.
2. Полученные результаты говорят о роли категориального сходства в возникновении феномена ПППП, а, следовательно, свидетельствуют в пользу основных положений теории установки в отношении работы механизмов восприятия и внимания и с наибольшей полнотой могут быть объяснены в рамках комбинированной теории, сочетающей в себе положения теории установки и теории истощения ресурсов.
Литература
- Горбунова Е.С. Пространственная рабочая память при решении задачи зрительного поиска множественных стимулов // Экспериментальная психология. 2017. Т. 10. № 1. С. 38—52.
- Adamo S.H., Cain M.S., Mitroff S.R. Self-Induced Attentional Blink: A Cause of Errors in Multiple- Target Search // Psychological Science. 2013. Vol. 24(12). P. 2569—2574. doi: 10.1177/0956797613497970
- Adamo S.H., Cain M.S., Mitroff S.R. Targets need their own personal space: effects of clutter on multiple-target search accuracy // Perception. 2015. Vol. 44(10). P. 1203—1214. doi: 10.1080/13506285.2012.726448
- Berbaum K.S., Franken Jr.E.A., Dorfman D.D., Rooholamini S.A., Coffman C.E., Cornell S.H., Cragg A.H., Galvin J.R., Honda H., Kao S.C., et al. Time course of satisfaction of search // Investigative Radiology. 1991. Vol. 26 (7). P. 640—648.
- Biggs A.T., Adamo S.H., Dowd E.W., Mitroff S.R. Examining perceptual and conceptual set biases in multiple-target visual search // Attention, Perception, Psychophysics. 2015. Vol. 77(3). P. 844—855. doi: 10.3758/s13414-014-0822-0
- Cain M.S., Mitroff S.R. Memory for found targets interferes with subsequent performance in multiple-target visual search // The Journal of Experimental Psychology: Human Perception and Performance. 2013. Vol. 39(5). P. 1398—1408. doi: 10.1037/a0030726
- Cain M.S., Biggs A.T., Darling E.F., Mitroff S.R. A little bit of history repeating: Splitting up multiple-target visual searches decreases second-target miss errors // Journal of Experimental Psychology: Applied. 2014. Vol. 20(2). P. 112—125. doi: 10.1037/xap0000014
- Duncan J., Humphreys G.W. Visual search and stimulus similarity // Psychological Review. 1989. Vol. 96 (3). P. 433—458.
- Fleck M.S., Samei E., Mitroff S.R. Generalized “Satisfaction of Search”: Adverse Influences on Dual- Target Search Accuracy // Journal of Experimental Psychology. Applied. 2010. Vol. 16(1). P. 60—71. doi: 10.1037/a0018629
- Gorbunova E. Perceptual similarity in visual search for multiple targets // Acta Psychologica. 2017. Vol. 173. P. 46—54. doi: 10.1016/j.actpsy.2016.11.010
- Kristjánsson Á., Campana G. Where perception meets memory: A review of repetition priming in visual search tasks // Attention, Perception, & Psychophysics. 2017. Vol. 72. P. 5—18. doi: 10.3758/APP.72.1.5
- Kwak H.W., Dagenbach D., Egeth H. Further evidence for a time-independent shift of the focus of attention // Perception & Psychophysics. 1991. Vol. 49 (5). P. 473—480.
- Schwarz W., Eiselt A.K. Numerical distance effects in visual search // Attention, Perception, & Psychophysics. 2012. Vol. 74(6). P. 1098—1103. doi: 10.3758/s13414-012-0342-8
- Tuddenham W.J. Visual search, image organization, and reader error in roentgen diagnosis // Radiology. 1962. Vol. 78. P. 694—704.
- Visser T.A., Davis C., Ohan J.L. When similarity leads to sparing: probing mechanisms underlying the attentional blink // Psychological Research. 2009. Vol. 73 (3). P. 327—335. 10.1007/s00426-008-0155-5
Информация об авторах
Метрики
Просмотров web
За все время: 1831
В прошлом месяце: 31
В текущем месяце: 23
Скачиваний PDF
За все время: 750
В прошлом месяце: 8
В текущем месяце: 2
Всего
За все время: 2581
В прошлом месяце: 39
В текущем месяце: 25